При каких условиях происходит оплодотворение. Оплодотворение. Искусственное оплодотворение яйцеклетки
Созревание ооцитов начинается еще до рождения ребенка. Первичные половые клетки, мигрировав в гонаду женщины, путем дифференциации превращаются в оогонии.
Оогенез
Оогонии совершают многочисленные митозы и в конце третьего месяца эмбрионального развития формируют кластеры, окруженные одним слоем плоских эпителиальных клеток (фолликулярных клеток, происходящих из поверхностного эпителия яичника) и образуют примордиальные фолликулы.
Оба этих прямых и более косвенных исследования указывают на значительную обратную зависимость между средним размером яиц у женщин и количеством спермы, необходимой для оплодотворения 50% яиц. Эти исследования показывают, что женщины значительно различаются по размерам яиц, которые они производят, и что эти различия в размерах коррелируют с вероятностью оплодотворения.
Не каждое исследование подтвердило, что женщины, производящие большие яйца, получают более высокие показатели оплодотворения. Затем яйца от этих самок собирали и объединяли в две группы обработки, а группа с низким содержанием пищи имела значительно более крупные яйца и проявляла заметную тенденцию к уменьшению оплодотворения при воздействии на сперму с одной и той же низкопродуктивной обработкой. Поскольку анализ оплодотворения не был реплицирован, яйца подвергались воздействию только одной и неизвестной концентрации спермы, а яйца от обработки с низким содержанием пищи подвергались только воздействию спермы из той же самой обработки; интерпретировать эти результаты непросто.
Оогонии продолжают митотическое деление, а некоторые из них подлежат большей дифференциации и образуют первичные ооциты. Сразу после образования первичные ооциты реплицируют свою ДНК и вступают в профазу первого мейотического деления. В течение следующих месяцев количество оогоний быстро растет, и к пятому месяцу развития общее количество половых клеток в яичнике достигает максимума — около 7 млн.
Для этой тенденции существует несколько возможностей. Во-первых, голодные самцы, возможно, произвели меньше спермы. Во-вторых, голодные самки могли производить яйца низкого качества. В-третьих, и, что менее вероятно, большие яйца от лечения с низким содержанием пищи могли бы иметь повышенные уровни полиспермии.
Независимо от объяснения, это исследование действительно предполагает, что простая мера размера яйца, независимо от других факторов, не является хорошим предиктором успеха оплодотворения. Не все изменения в успехе оплодотворения могут быть объяснены различиями в размере яиц у женщин. Другие факторы, такие как аксессуарные структуры яиц и внутривидовые различия в гаметной совместимости или другие аспекты качества гамет, также могут способствовать изменению успеха оплодотворения.
В это время начинается массовая гибель клеток и многочисленные оогонии, как и первичные ооциты, испытывают атрезии. К седьмому месяцу эмбрионального развития большинство оогоний дегенерирует, за исключением небольшого их количества вблизи поверхности яичника. Все первичные ооциты окружены одним слоем плоских эпителиальных клеток и формируют примордиальные фолликулы.
Одним из способов уменьшить генетические различия между яйцами, которые могут способствовать различиям в совместимости или другим генетическим или материнским влияниям на размер яйца, является изучение фенотипической вариации размера яйца у одной женщины. Размер яиц варьируется у самки, и этот вариант коррелирует с успехом оплодотворения. Испытания, которые сравнивают размеры яиц, высвобождаемых самкой перед добавлением спермы, и размеры яиц, которые остаются неоплодотворенными после добавления предельного количества спермы, так что оплодотворяют только часть яиц, указывают, что более крупные яйца предпочтительно оплодотворяются, когда сперма ограничивается.
Перед рождением ребенка все первичные ооциты уже вступили в профазу мейоза, но вместо перехода к метафазе они вступают в стадию диплотена (специфическую стадию покоя во время профазы), которая характеризуется образованием кружевной сетки хроматина. Первичные ооциты остаются в профазе и не завершают первого мейотического деления вплоть до достижения пубертатного периода (периода ). Это, возможно, происходит под действием ингибитора созревания ооцита, который образуется фолликулярными клетками.
В этих исследованиях, когда добавляли более высокие концентрации спермы, все яйца оплодотворялись. Поэтому мелкие яйца не являются неоплодотворяемыми, их просто менее вероятно оплодотворяют в условиях ограничения спермы. Эти тесты включают эхиноиды, асцидии и полихеты. Кроме того, Маршалл и его коллеги отметили каскадные эффекты условий ограничения спермы: большие яйца оплодотворяются, приводя к большим эмбрионам и личинкам, более высокому вылупляющему успеху и более коротким временам развития, чем яйца, оплодотворенные в условиях с обильной спермой.
Считают, что общее количество первичных ооцитов при рождении девочки колеблется от 700 000 до 2 млн. Течение последующих лет жизни большинство ооцитов подвергается атрезии, и только около 400 000 клеток остаются до начала пубертатного периода, из которых менее 500 подвергаются овуляции в репродуктивном периоде. Важным является тот факт, что некоторые ооциты, которые достигают зрелости позже, находятся в состоянии покоя (в диплотенний фазе первого мейотического деления) в течение 40 и более лет.
Одна потенциальная проблема смешения, связанная с использованием искусственно порожденных яиц у одиноких самок, заключается в том, что возможно, что более мелкие яйца развиты менее зрелыми по сравнению с более крупными яйцами. Хотя часто предпринимаются попытки использовать зрелых индивидуумов и гамет, и исследования обычно показывают, что подавляющее большинство яиц являются удобряемыми с учетом оптимальной концентрации спермы, могут быть тонкие различия в развитии среди яиц, которые коррелируют с размером яйца.
Самый сильный случай гипотезы о том, что размер яйца влияет на успех оплодотворения, заключается в том, что этот эффект прогнозируется с помощью моделей оплодотворения-кинетики, и эта картина отмечена на разных организационных уровнях. Проблемы, отмеченные на одном уровне, менее вероятно, будут проблемой на других уровнях. Хотя многие факторы качества гаметы, вероятно, влияют на оплодотворение, эффект размера яйца обычно поднимается выше этих источников необъяснимой дисперсии до значимости.
До сих пор неизвестно, есть ли диплотеновая стадия наиболее благоприятной для защиты ооцита от агрессивных факторов внешней среды, которые действуют на яичник на протяжении всей жизни женщины. Доказано, что риск рождения детей с хромосомными аномалиями увеличивается с возрастом матери: старения первичных ооцитов делает их уязвимыми к повреждениям.
Как вспомогательные структуры влияют на успех оплодотворения?
Размер мишени, влияющий на столкновения сперматозоидов, может зависеть от вспомогательных структур, которые могут увеличить эффективный размер яйца. Эти вспомогательные структуры включают в себя ряд внеклеточных структур, обнаруженных на различных таксонах. Кроме того, химикаты, выделяемые яйцами, могут привлекать сперму или стимулировать их активность.
Наиболее изученной аксессуарной структурой является желе, покрывающее яйца эхиноидов. Желевые пальто были вовлечены в ряд процессов, важных для пригодности яйца, включая защиту от сдвиговых сил, таких как выброс яйца у самки, увеличение уровней активности спермы, подготовка спермы для оплодотворения, предоставление видовой специфичности, обеспечение барьера к полиспермии и увеличению эффективного целевого размера яйца.
При достижении половой зрелости каждый очередной овариальный цикл начинается со вступления в фазу созревания 5-15 примордиальных фолликулов. Первичный ооцит, который находится в стадии диплотена, начинает расти, а окружающие его фолликулярные клетки превращаются из плоских в кубические, пролиферируют и образуют многослойный эпителий зернистой зоны (гранулезы), таким образом образуется первичный фолликул. Клетки гранулезы отделены базальной мембраной от окружающих стромальных клеток, которые образуют внешнюю оболочку, крышу — папку фолликула.
Работа над тем, как желеобразные пальто может повлиять на оплодотворение, началась в начале прошлого века. Эксперименты показали, что зачистка яиц эхиноидов их желеобразных покрытий снижает процент оплодотворенных яиц, но повышает скорость оплодотворения. Этот результат согласуется с представлением о том, что желеобразные покрытия повышают скорость столкновений, но это потому, что для проникновения спермы в этот слой требуется время, скорость оплодотворения уменьшается.
Аналогичные результаты были отмечены в более поздних исследованиях. Как и было предсказано, коллизии чаще наблюдались при более высоких концентрациях спермы и с яйцами с желеобразными пальто. Когда частота столкновений регулировалась по площади поверхности, столкновения с пластиковыми шариками и те, у которых яйца с и без желеобразных покрытий, существенно не различались. Этот результат позволяет предположить, что частоты столкновений могут быть объяснены у этого вида по размеру мишени. Оценки оплодотворения, сделанные за короткий промежуток времени, показали, что желеобразные покрытия повышают вероятность оплодотворения, но не до степени, предсказанной увеличением целевого размера.
Клетки гранулезы и ооцит секретируют гликопротеины, которые образуют на поверхности ооцита прозрачную зону. В течение роста фолликула клетки наружной оболочки разделяются на два слоя: внутренний слой секреторных клеток — внутреннюю папку и внешний слой соединительной ткани, содержащей фибробластоподобные клетки — внешнюю папку. Фолликулярные клетки образуют маленькие пальцеобразные отростки, проходящие через прозрачную зону и могут играть важную роль в транспортировке веществ из фолликулярных клеток к .
Этот результат может возникнуть, потому что сперматозоиды, сталкивающиеся с желеобразным слоем, могут быть менее склонны оплодотворять яйцо по сравнению с спермой, сталкивающейся непосредственно с поверхностью яйца. Последний эксперимент в этой серии был предназначен для более глубокого отражения естественных условий. Мужские и женские морские ежи из одного из трех конъюнктов морского ежа были вызваны в поле при естественных плотностях. После 30 мин нереста яйца были собраны над каждой нерестовой самкой, и было зарегистрировано распределение и численность всех морских и морских млекопитающих.
В течение оогенеза между клетками гранулезы появляются заполненные жидкостью пространства, при слиянии которых образуется пещера (антрум), и фолликул становится вторичным. Сначала пещера имеет форму полумесяца, но затем увеличивается. Клетки гранулезы, локализующиеся вокруг ооцита, формируют яйценосный бугорок. Зрелый фолликул, который имеет диаметр> 10 мм, называется третичным или пузырчатым, граафовым фолликулом. Третичный фолликул окружен внутренней папкой, которая образована стероидсекретирующими клетками и богатая кровеносные сосуды, и внешней папкой, которая постепенно переходит в строму яичника.
Эксперимент повторялся 20 раз для каждого вида. Однако при анализе ковариации, которая скорректировала средний успех оплодотворения на основе средних значений ковариатов, порядок рангов ответа был отменен; у видов с самым высоким уровнем оплодотворения были наихудшие показатели оплодотворения, а виды с самым низким уровнем оплодотворения имели лучшие показатели оплодотворения при средних значениях ковариации. Этот результат был подтвержден экспериментальными манипуляциями с плотностью населения. Эти результаты согласуются с представлением о том, что видоспецифические гаметные признаки адаптированы к типичным уровням доступности сперматозоидов для каждого вида и что различия в размерах яиц, по крайней мере, коррелируют с этими образцами.
В каждом яичниковом цикле в стадию роста вступают несколько фолликулов, но обычно лишь один из них достигает полной зрелости, тогда как остальные подлежат дегенерации (атрезии). После достижения зрелости фолликула первичный ооцит в нем восстанавливает первое митотическое деление (мейоз I), что приводит к образованию двух неодинаковых по размерам дочерних клеток, каждая из которых имеет 23 двохроматидные хромосомы.
Интересно отметить, как эти гаметные черты и характеристики оплодотворения различаются в разных регионах, но, как и при любой адаптивной признаке, попытка сопоставить значения признаков с существующими экологическими условиями требует предположения о том, что нынешние условия сохраняются достаточно долго, чтобы влиять на черты, о которых идет речь.
Исторические плотности многих видов, включая этих морских ежей, возможно, были совершенно разными в прошлые времена, что сделало это предположение сомнительным. Поведенческая склонность этих видов к скоплению или диспергированию может быть более надежным анализом исторических оценок местных условий спермы, чем более масштабные оценки численности. Таким образом, эмпирические результаты предполагают, во-первых, что размер яйца коррелирует с успехом оплодотворения в лаборатории внутри самки, у самок внутри видов и часто по видам.
Одна из этих клеток, вторичный ооцит, получает большую часть цитоплазмы, а вторая, первое полярное тельце, цитоплазмы почти не имеет. Первое полярное тельце локализуется между прозрачной зоной и клеточной оболочкой вторичного ооцита в перивителиновом пространстве. Первое мейотическое разделение восстанавливается непосредственно перед овуляцией.
Во-вторых, этот результат не скрыт у видов, которые, как было показано, имеют вспомогательные структуры или хемотаксис спермы, которые могут повлиять на успех оплодотворения. Наконец, полевые характеристики гамет в разных условиях потока чувствительны к изменению яичных признаков внутри и среди видов, и эта вариация коррелирует с размером яйца.
Предполагают ли теоретические модели, что доступность спермы может влиять на эволюцию размера яйца?
Эмпирические данные свидетельствуют о том, что крупные яйца оплодотворяются более высокими темпами в условиях ограничения спермы. Чтобы определить, способствует ли селекция увеличению размера яйца, стоит учитывать все последствия изменения размера яйца, включая как преимущества, так и затраты на увеличение размера яйца. Первые включают сокращение времени разработки или увеличение выживаемости; последние включают дополнительные ресурсы, которые должны быть выделены на потомство, что влечет за собой либо увеличение общего распределения на воспроизводство, либо уменьшение числа потомков.
После завершения первого мейотического деления вторичный ооцит, не возвращаясь в стадию покоя и не реплицируя ДНК, вступает во второе мейотическое деление (мейоз II). Во время образования во вторичном ооците митотического веретена и метафазной хромосомной пластинки происходит овуляция. Под давлением накопившейся фолликулярной жидкости истонченный участок стенки граафового фолликула разрывается, и ооцит выталкивается из яичника в просвет маточной трубы.
Были построены различные модели оптимальности, которые учитывают различные затраты и выгоды от увеличения размера яйца и предполагают различные функциональные связи между размером яйца и фитнесом, и все они предполагают, что изменение доступности спермы может влиять на эволюцию размера яйца.
Сортировка прогнозируемых эффектов доступности спермы на размер яйца осложнялась различными предположениями, используемыми моделями для описания взаимосвязи между размером яйца и временем разработки. Это предположение имеет решающее значение, поскольку оно в значительной степени определяет, как фитнес-потомство относится к размеру потомства. Более ранние модели оптимального размера яйца морских беспозвоночных предполагали отрицательную линейную зависимость между размером яйца и временем развития, что приводит к предсказаниям экстремумов в размере яйца.
Второй раздел созревания завершается только в случае оплодотворения ооцита. Если оплодотворение не происходит, ооцит погибает (дегенерирует) через 24 ч после овуляции. Неизвестно, осуществляет первое полярное тельце второй раздел, хотя описаны случаи наблюдения оплодотворенной яйцеклетки в сопровождении трех полярных телец.
Сперматогенез
Созревание сперматозоидов (сперматогенез), в отличие от ооцитов, начинается только в пубертатном периоде и включает процессы преобразования сперматогония в сперматозоиды. При рождении мальчика половые клетки находятся в половых шнурах яичек (маленькие светлые клетки, окруженные поддерживающими клетками Сертоли). Поддерживающие клетки Сертоли происходят из клеток поверхностного эпителия яичка, подобно фолликулярных клеток яичника.
Вскоре перед половым созреванием в половых шнурах возникает просвет, и они превращаются в семенные канальцы. В настоящее время первичные половые клетки дают начало , среди которых есть 2 типа: сперматогоний типа А и сперматогоний типа Б. Сперматогонии типа А подразделяются митозом и обеспечивают резерв стволовых клеток, сперматогонии типа Б дают начало первичным сперматоцитам.
При нормальных условиях некоторые клетки типа А выходят из популяции стволовых клеток, чтобы начать дальнейшие генерации сперматогония, каждая из которых является более дифференцированной по сравнению с предыдущей генерацией. После завершения последнего деления клеток типа А формируются сперматогоний типа Б, который после серии митозов превращаются в первичные сперматоциты. Первичные сперматоциты вступают в продолженную профазу (22 дня), после которого наступает быстрое завершение мейоза І с образованием вторичных сперматоцитов.
Вторичные сперматоциты сразу начинают второе мейотическое разделение, в результате чего образуются гаплоидные сперматиды. В течение этих процессов, начиная с момента, когда клетки типа А оставляют популяцию стволовых клеток, и к образованию сперматида, цитокинез не заканчивается, и клетки последовательных генераций остаются связанными цитоплазматическими мостиками. Итак, потомки одного сперматогонического типа А формируют кластер половых клеток, сохраняющих контакт между собой на протяжении всей дифференциации.
Кроме того, сперматогоний и сперматиды протяжении своего развития погружены в глубокие карманы из клеток Сертоли. Клетки Сертоли обеспечивают поддержку для созревающих половых клеток, их защиту и питание, создают условия для высвобождения зрелых сперматозоидов.
Спермиогенез — ряд изменений, в результате которых сперматиды превращаются в сперматозоиды. У человека процесс преобразования сперматогоний в зрелый сперматозоид продолжается 64 суток. включает следующие фазы:
1) формирование акросомы, покрывающей половину поверхности ядра и содержащей энзимы, помогающие пенетрации в яйцеклетку через ее микроокружение во время оплодотворения;
2) конденсацию ядра;
3) формирование шейки, средней части и хвоста;
4) отторжение большей части цитоплазмы.
Сформированные сперматозоиды направляются в просвет семенных канальцев. С семенных канальцев путем сокращения элементов их стенки сперматозоиды продвигаются в канальцы придатка яичка. Сначала сперматозоиды малоподвижны и приобретают активную подвижность в придатке яичка.
Клинические корреляции. Аномальные гаметы . У человека, как и у большинства млекопитающих, один фолликул яичника иногда может содержать 2 или 3 первичных ооцита. Такие ооциты могут дать начало двойни или тройни, но обычно они дегенерируют, не достигнув стадии зрелости. В редких случаях один ооцит может содержать два или три ядра. Такие многоядерные ооциты также погибают, не достигнув зрелости.
В отличие от ооцитов, аномалии сперматозоидов встречаются часто (около 10% сперматозоидов имеют дефекты). Головка, хвост, сперматозоиды могут быть маленькими, гигантскими или соединенными. Сперматозоиды с отклонениями в морфологии не имеют нормальной подвижности и обычно не способны к оплодотворению ооцитов.
— это процесс слияния мужской и женской гамет. Оплодотворение происходит в ампулярной части маточной трубы.
Ооцит, который выталкивается из фолликула вместе с окружающими его клетками гранулезы участка яйценосного бугорка, попадает в ампулу маточной трубы. Некоторые клетки яйценосного бугорка после этого переориентируются вокруг прозрачной зоны и образуют лучистый венец, или корону. В момент выброса ооцита, окруженного клетками яйценосного бугорка, из яичника (овуляция) завершается первое мейотическое разделение и вторичный ооцит начинает второе мейотическое разделение.
Сперматозоиды сохраняют способность к оплодотворению в женских половых путях в течение нескольких суток. Они быстро проникают из влагалища в шейку матки, матку и маточные трубы, что обусловлено двигательной активностью хвостов сперматозоидов в шеечной слизи и сокращениями мускулатуры матки и маточной трубы. Для сперматозоидов, которые оказываются в маточной трубе, перешеек служит резервуаром, а движение с этого участка в ампулы является синхронизированным процессом.
Сперматозоиды, которые только попали в женские половые пути, не способны к оплодотворению ооцита. Для приобретения способности к оплодотворению им нужно осуществить капацитацию и акросомную реакцию.
Капацитация — это химическая модификация (кондиционирование) поверхности сперматозоидов и семенной жидкости в женских половых путях. У человека этот процесс длится около 7 часов. Суть капацитации, которая происходит в маточной трубе, состоит во взаимодействии между сперматозоидом и поверхностью слизистой оболочки маточной трубы . При этом гликопротеиновые покрытия и белки семенной жидкости удаляются из плазматической мембраны акросомного участка сперматозоида. Только капацитованнные сперматозоиды могут пройти сквозь лучистый венец ооцита и осуществить акросомную реакцию.
Акросомная реакция происходит после связывания сперматозоида с прозрачной зоной и обусловлена белками последней. Суть акросомной реакции заключается в высвобождении энзимов, необходимых для пенетрации прозрачной зоны — акрозина и трипсиноподобных веществ.
состоит из трех фаз, или стадий:
1) пенетрация лучистого венца;
2) пенетрация прозрачной зоны;
3) слияние мембран ооцита и сперматозоида.
В стадии 1 из 200-300 млн сперматозоидов, которые попадают в женские половые пути при семяизвержении, только 300-500 достигают места оплодотворения и только один из них оплодотворяет яйцеклетку; другие сперматозоиды, возможно, помогают ему проникнуть сквозь барьеры, окружающие ооцит. Капацитованные сперматозоиды легко проникают через клетки лучистого венца.
В стадии 2 прозрачная зона— гликопротеиновая оболочка, окружающая ооцит, обеспечивает связывание сперматозоидов и индуцирует акросомную реакцию. Связывание сперматозоидов с прозрачной зоной индуцируется лигандом 2Р3, белком прозрачной зоны и рецепторами сперматозоида. Высвобождение акросомных энзимов (акрозина) позволяет сперматозоиду проникнуть сквозь прозрачную зону и вступить в контакт с плазматической мембраной ооцита.
После взаимодействия головки сперматозоида с поверхностью ооцита проницаемость прозрачной зоны меняется. Результатом этого взаимодействия является секреция лизосомных ферментов кортикальных гранул, содержащихся под плазмолеммой ооцита. Эти ферменты вызывают изменение свойств прозрачной зоны, что предотвращает пенетрации других сперматозоидов и инактивирует видоспецифические сайты рецепторов сперматозоидов на поверхности прозрачной зоны. В прозрачную зону могут углубляться много сперматозоидов, но лишь один из них проникает в ооцит.
В стадии 3 первичная адгезия сперматозоида с ооцитом частично опосредуется взаимодействием интегринов поверхности ооцита и их лигандов на сперматозоиды. Сразу после адгезии происходит слияние цитоплазматических мембран сперматозоида и яйцеклетки. Поскольку плазматическая мембрана акросомного участка сперматозоида исчезает в результате акросомной реакции, сливаются мембрана ооцита и мембрана задней части головки сперматозоида. У человека в цитоплазму ооцита входят головка и хвост сперматозоида, тогда как плазмолемма сперматозоида остается фиксированной к поверхности ооцита.
На проникновение сперматозоида ооцит реагирует тремя процессами:
1. Кортикальная реакция и зона-реакция. В результате выброса из ооцита кортикальных гранул, в которых содержатся лизосомальные ферменты, мембрана ооцита становится непроницаемой для других сперматозоидов и прозрачная зона меняет свою структуру и состав, предотвращая связывание и проникновению других сперматозоидов. Этот механизм предотвращает полиспермию (проникновение в ооцит более чем одного сперматозоида).
2. Обновление второго мейотического деления. Сразу после вхождения сперматозоида ооцит завершает свой второй мейотическое разделение. Одна из дочерних клеток почти не получает цитоплазмы — второе полярное тельце, другая — является дефинитивным ооцитом. Хромосомы дефинитивных ооцита (22 + Х) образуют пузырчатое ядро — женский .
3. Метаболическая активация яйцеклетки. Фактор активации, возможно, вносится сперматозоидом. Активация после слияния ооцита со сперматозоидом включает в себя первичные клеточные и молекулярные процессы, связанные с ранним эмбриогенезом.
После проникновения через оболочки ооцита сперматозоид движется вперед, к достижению тесной близости с женским пронуклеусом. Ядро сперматозоида набухает и образует мужской пронуклеус, тогда как хвост отделяется и дегенерирует.
Морфологически мужской и женский пронуклеусы различить невозможно. Они вступают в тесный контакт, теряют ядерные оболочки и реплицируют свои ДНК. Сразу после синтеза ДНК хромосомы образуют веретено деления и готовятся к митозу. Материнские (23) и родительские (23) двохроматидные хромосомы расщепляются продольно в области центромеры. Сестринские хроматиды движутся к противоположным полюсам, обеспечивая каждую клетку двухклеточного зародыша нормальным диплоидным количеством хромосом и ДНК. Когда сестринские хроматиды движутся к противоположным полюсам, на поверхности клетки образуется глубокая борозда, которая со временем разделяет цитоплазму на 2 части.
Итак, основные результаты оплодотворения включают следующие процессы:
1) восстановление диплоидного набора хромосом, половина из которых происходит от отца, половина — от матери. Образованная зигота содержит новую комбинацию , отличается от генотипа родителей;
2) определение пола нового индивида. Сперматозоид, несущий Х-хромосому, обусловливает развитие эмбриона женского пола (ХХ), а сперматозоид, несущий У-хромосому, эмбриона мужского пола (ХУ);
3) начало дробления. Если оплодотворение не происходит, ооцит обычно дегенерирует через 24 ч после овуляции.
Клинические корреляции . Проблема бесплодия затрагивает 15-30% супружеских пар. Мужское бесплодие обусловлено аномалиями количества и качества сперматозоидов. В норме эякулят имеет объем 3-4 мл и содержит 100 млн сперматозоидов. Если количество сперматозоидов <50 млн в эякуляте (или <20 млн в 1 мл спермы), имеет место олигозооспермия и мужская бесплодие.
Женское бесплодие может быть обусловлено анатомическими факторами (непроходимость маточных труб и др.), отсутствием овуляции (ановуляцией), неблагоприятным влиянием цервикальной слизи на сперматозоиды; иммунитетом к сперматозоидам и другими причинами.
При экстракорпоральном оплодотворении, или фертилизации , рост фолликулов в яичниках стимулируют введением гонадотропинов; забирают ооциты перед овуляцией, помещают их в культуральную среду и добавляют сперматозоиды. Оплодотворенные яйцеклетки культивируют до 8-клеточной стадии, а затем имплантируют в матку. Эмбрионы доимплантационных стадий нечувствительны к действию тератогенных факторов, поэтому риск пороков развития при осуществлении манипуляций при экстракорпоральном оплодотворении не увеличивается.
Дробление
После достижения зародышем двухклеточной стадии он осуществляет серию митотических делений (процесс дробления), что приводит к увеличению количества клеток. Эти клетки (бластомеры) уменьшаются с каждым делением дробления и к восьмиклеточной стадии формируют неплотный конгломерат. После третьего раздела дробления бластомеры максимально увеличивают площадь межклеточных контактов, формируя компактную клеточную массу, которая содержится вместе благодаря плотным контактам.
Процесс компактизации приводит к отделению внешних клеток от внутренних, которые сочетаются между собой щелевыми контактами (Нексус). Примерно через 3 ч после оплодотворения клетки компактизированного эмбриона снова делятся, образуя 16-клеточную морулы (ягода шелковицы). Внутренние клетки морулы образуют внутреннюю клеточную массу, а наружные клетки — наружную клеточную массу. Внутренняя клеточная масса дает начало тканям эмбриона, а внешняя клеточная масса образует трофобласт, из которого формируется .
Бластоциста
Когда морула попадает в полость матки, через прозрачную зону с микроокружения эмбриона начинает просачиваться жидкость, которая накапливается в межклеточных пространствах внутренней клеточной массы. Постепенно межклеточные пространства сливаются и формируют единую полость — бластоцель. Клетки внутренней клеточной массы образуют эмбриобласт и локализованы на одном полюсе, тогда как клетки наружной клеточной массы или трофобласт, становятся плоскими и образуют стенку бластоцисты. Прозрачная зона исчезает, что дает возможность бластоцистам осуществить имплантацию.
Клетки трофобласта, которые локализуются над эмбриобластным полюсом бластоцисты, около шестого дня начинают врастать в эпителий эндометрия — слизистой оболочки матки. Прикрепление и инвазия трофобласта обеспечиваются действием интегринов, выделяемых клетками трофобласта, а также ламинином и фибронектином межклеточного матрикса эндометрия.
Имплантация является результатом комплексных взаимодействий трофобласта и эндометрия. К концу первой недели развития зародыш человека проходит стадию морулы, бластоцисты и начинает имплантацию в слизистую оболочку матки. Итак, имплантация происходит в конце первой недели развития.
Клинические корреляции . Аномальные зародыши обычно погибают через 2-3 нед после оплодотворения, поэтому их частоту определить трудно. Полагают, что около 50% беременностей заканчиваются самопроизвольными выкидышами, и половина из них обусловлена хромосомными аномалиями. Эти выкидыши является следствием так называемых «пренатальных фильтров», что отбраковывают аномальные эмбрионы и тем самым уменьшают количество индивидов с врожденными пороками до 2-3% вместо 12%.
При комбинации методов экстракорпорального оплодотворения и полимеразной цепной реакции (ПЦР) проводится молекулярный скрининг эмбрионов с генетическими аномалиями (преимплантационная диагностика). Для этого достаточно выделить один бластомер из эмбриона ранней стадии развития и реплификовать его ДНК для дальнейшего исследования. Программа «Геном человека» занимается изучением связи специфических генов с различными клиническими синдромами.
Двухслойный зародышевый диск
На второй неделе развития (8-е сутки) бластоциста частично погружена в строму эндометрия. В области над эмбриобластом трофобласт дифференцируется на два слоя: внутренний слой одноядерных клеток — внешнюю многоядерных массу без четких клеточных границ — синцитиотрофобласт.
Фигуры митоза наблюдаются только в цитотрофобласте; в синцитиотрофобласте они отсутствуют. Клетки цитотрофобласта делятся и мигрируют в синцитиотрофобласт, там сливаются и теряют свои индивидуальные клеточные оболочки. Клетки эмбриобласта также дифференцируются на два слоя: слой маленьких кубоидальних клеток, прилегающих к полости бластоцисты, — гипобласт, и слой высоких цилиндрических клеток, прилегающих к амниотической полости, — эпибласт. Вместе эти два слоя эмбриобласта образуют плоский двухслойный зародышевый диск.
Одновременно внутри эпибласта образуется маленькая полость, которая со временем расширяется и образует амниотическую полость. Клетки эпибласта, прилегающие к цитотрофобласте (амниобласты), вместе с другими клетками эпибласта выстилают амниотическую полость. Строма эндометрия вокруг места имплантации характеризуется отеком и выраженной васкуляризацией; большие разветвленные железы продуцируют значительное количество гликогена и слизи.
На 9-10-е сутки бластоциста глубже погружается в эндометрий. Трофобласт на эмбриональном полюсе развивается наиболее интенсивно, в его синцитии образуются вакуоли, которые со временем сливаются в большие лакуны — лакунарная стадия развития трофобласта. Этим временем на противоположном полюсе бластоцисты плоские клетки, возможно, гипобластического происхождения, формируют тонкую экзоцеломическую мембрану — мембрану Гейзера, которая выстилает внутреннюю поверхность цитотрофобласта. Мембрана Гейзера вместе с гипобластами образует выстилки экзоцеломической полости — первичного желточного мешка.
На 11-12-е сутки развития бластоциста полностью погружается в строму эндометрия, а поверхностный эпителий закрывает первичный дефект эндометрия. Бластоциста несколько выпячивается в просвет матки. образует лакунарные пространства в синцитии, которые сочетаются между собой с образованием сетки. Эта сетка наиболее выражена на эмбриональном полюсе, тогда как на противоположном полюсе трофобласт образован преимущественно клетками цитотрофобласта. Клетки синцитиотрофобласта проникают в строму эндометрия и разрушают едотелиальний слой материнских капилляров. Эти капилляры становятся расширенными, синусоидными.
Синцитиальная лакуны сочетаются с синусоидными капиллярами, и материнская кровь попадает в лакунарную систему трофобласта. При разрушении трофобластом все большего количества синусоидов материнская кровь начинает циркулировать через лакунарную систему, возникает маточно-плацентарный кровоток. Между внутренней поверхностью цитотрофобласта и внешней поверхностью экзоцеломической полости появляется новая популяция клеток. Эти клетки происходят из желточного мешка и образуют рыхлую соединительную ткань — внезародышевую мезодерму, которая заполняет все пространства между трофобластом снаружи и амнионом и экзоцеломичсекой мембраной изнутри.
Вскоре в внезародышевой мезодерме возникают большие полости, при слиянии которых образуется пространство — полость хориона. Это пространство окружает первичный желточный мешок и амниотическую полость, за исключением того участка, где зародышевый диск прикрепляется к трофобласту соединительной ножкой. Та часть внезародышевой мезодермы, которая выстилает цитотрофобласт и амнион, называется внезародышевой соматоплевральной мезодермой. А часть, покрывающая желточный мешок, называется внезародышевой спланхноплевральной мезодермой.
Рост двухслойного зародышевого диска происходит относительно медленно. Клетки эндометрия в это время становятся полигональными, накапливают гликоген и липиды, межклеточные пространства накапливают жидкость, ткань становится отечной — происходит децидуальная реакция, сначала охватывающая место имплантации, а затем распространяющаясяя на весь .
К 13-м суткам дефект на поверхности эндометрия заживает, но иногда с места имплантации может возникать кровотечение вследствие увеличения кровотока в лакунарных пространствах. Это кровотечение происходит около 28-го дня цикла и может восприниматься как менструация, что приводит к неточному определению гестационного возраста.
Клетки цитотрофобласта пролиферируют и врастают в синцитиотрофобласт, формируя окруженные синцитием клеточные колонки — первичные ворсинки. Из гипобласта тогда выселяются клетки, которые мигрируют вдоль внутренней поверхности экзоцеломической мембраны, пролиферируют и формируют новую полость внутри экзоцеломической полости — вторичный, или дефинитивный желточный мешок. Этот желточный мешок значительно меньше, чем первичная экзоцеломическая полость, или первичный желточный мешок.
При формировании дефинитивного желточного мешка большие части экзоцеломической полости сжимаются и из них образуются экзоцеломические пузырьки во внезародышевом целоме — хорионической полости. Внезародышевая мезодерма, что выстилает внутри цитотрофобласт, получает название хорионической пластинки. Единственный участок, где внезародышевая мезодерма пронизывает хорионическую полость, называется соединительной ножкой, которая с развитием кровеносных сосудов превращается в пупочный канатик.
В конце второй недели развития зародышевый диск представлен двумя прилегающими друг к другу клеточными дисками: эпибласт, который формирует дно амниотической полости, продолжает расширяться, и гипобласт, который формирует крышу вторичного желточного мешка. В главной области гипобласт образует утолщение из цилиндрических клеток, которые плотно прилегают к диску эпибласта — щечно-глоточная перепонка.
Клинические корреляции. Имплантация является критическим периодом развития эмбриона. Синцитиотрофобласта обеспечивает секрецию гормонов, в том числе хорионического гонадотропина человека (ХГЧ), определение которого в крови или моче лежит в основе тестов на беременность. Поскольку 50% генов имплантированного эмбриона имеют отцовское происхождение, он получужеродный для материнского организма.
Но отторжения зародыша не происходит благодаря защите комплекса факторов, которые вызывают продукцию цитокинов и протеинов, а также специфических молекул Н ^ А-С, принадлежащих к классу основного комплекса гистосовместимости и блокируют распознавания зародыша как посторонней ткани. При наличии у матери аутоиммунных заболеваний ( , антифосфолипидный синдром) отторжение эмбриона происходит за счет аутоантител, которые вторично атакуют зародыш.
Аномальная имплантация может иметь место даже в матке — не в передней или задней стенке, как обычно, а у внутреннего зева (предлежание плаценты). Имплантация вне матки приводит к развитию внематочной или эктопической беременности — в маточной трубе (95% случаев), в яичнике, брюшной полости.
Аномальные бластоцисты случаются очень часто. Большинство аномальных бластоцист не ставят проявлений беременности, потому что их трофобласт не способен поддерживать функцию желтого тела. Такие эмбрионы, вероятно, отторгаются с очередной менструацией, и поэтому беременность не наступает. В некоторых случаях аномальное развитие трофобласта приводит к развитию плацентарных оболочек при отсутствии или уменьшении эмбриональной ткани — и образуется пузырный занос, один из вариантов гестационной трофобластической болезни.
Другим вариантом этой болезни является развитие злокачественной клетки из тканей трофобласта - хориокарцинома . В случае образования пузырного заноса его клетки являются диплоидными, но полностью имеют отцовское происхождение, то есть имеет место оплодотворения ооцита, в котором отсутствует ядро, с последующей дупликацией мужских хромосом для восстановления диплоидного набора. Эти данные также свидетельствуют о том, что именно отцовские гены обусловливают развитие трофобласта, поскольку при пузырном заносе трофобласта формируется даже при отсутствии женского пронуклеуса.
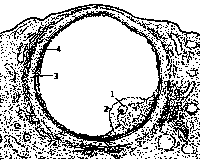
Менструальные циклы, подготовляющие организм к беременности, совершаются под влиянием гонадотропных гормонов гипофиза: фолликулостимулирующего (ФСГ - FSH), лютеинизирующего (ЛГ -LH) и лютеотропного (ЛТГ-LTH).Особо важная роль в регуляции циклических процессов принадлежит вегетативным центрам гипоталамуса, в которых образуется релизинг-гормон (RH), активирующий синтез и выделение гонадотропных гормонов гипофиза (ФСГ, ЛГ, ЛТГ).Яичниковый цикл. Под влиянием гонадотропных гормонов гипофиза в яичнике совершаются ритмически повторяющиеся изменения. Эти изменения сводятся к следующим фазам: а) развитие фолликула и разрыв созревшего фолликула - фолликулярная фаза; б) развитие желтого тела- лютеиновая (прогестероновая) фаза.Развитие фолликула и разрыв созревшего фолликула (фолликулярная фаза). Первичный (примордиальный) фолликул состоит из незрелой яйцеклетки, которая окружена одним слоем эпителиальных клеток (эпителий фолликула); снаружи фолликул окружен соединительнотканной оболочкой. Примордиальные фолликулы образуются в период внутриутробной жизни и раннего детства. К периоду половой зрелости из 400 000-500 000 первичных фолликулов остается 35 000-40 000, остальные подвергаются обратному развитию. Из оставшихся 35 000-40000 первичных фолликулов полностью созревают только 450-500, остальные подвергаются физиологической атрезии (фолликул начинает расти, но погибает, не достигнув полного развития).В первой - фолликулярной фазе менструального цикла начинается рост одного или нескольких фолликулов, но стадии полного созревания достигает обычно один фолликул. Другие фолликулы, рост которых начался вместе с нормально развивающимися, подвергаются атрезии и обратному развитию.Процесс созревания фолликула занимает первую половину менструального цикла, т. е. при 28-дневном цикле укладывается в 14 дней, а при 21-дневном - в 10-11 дней. В процессе развития фолликула существенные изменения претерпевают все его составные части; яйцеклетка, эпителий фолликула, соединительнотканная оболочка.Яйцеклетка увеличивается в размере в 5-6 раз, на ее поверхности образуется прозрачная оболочка (zona peleucida), в цитоплазме происходят сложные изменения. Окончательно яйцеклетка созревает после двукратного деления. Первое деление - созревательное; при нем образуются две неравные клетки. Основная, большая, клетка остается на месте (маленькая выбрасывается) и вскоре подвергается второму делению, при котором из клетки выбрасывается половина хромосом. После второго деления, которое называется редукционным, яйцеклетка становится зрелой и пригодной для оплодотворения (рис. 1).
| Фолликулярный эпителий в процессе созревания подвергается пролиферации- из однослойного становится многослойным, превращается в так называемую зернистую оболочку фолликула (membrana granulosa folliculi). Вначале клетки зернистой оболочки сплошной массой заполняют все пространство между яйцеклеткой и соединительнотканной оболочкой. Затем в центре скопления клеток образуются полости, заполненные жидкостью; впоследствии они сливаются и образуют одну общую полость, заполненную прозрачной фолликулярной жидкостью (liquor folliculi). По мере накопления фолликулярной жидкости клетки зернистой оболочки с одной стороны отодвигаются к яйцу, с другой - к стенке фолликула. Клетки зернистой оболочки, окружающие яйцеклетку, носят название лучистого венца (corona radiata). |
 |
Под его влиянием происходят: а) гиперплазия мышечных элементов матки в период полового созревания; б) пролиферация функционального слоя эндометрия в первой фазе менструального цикла; в) пролиферация выводных протоков молочной железы; г) размножение клеток эпителиального покрова слизистой оболочки влагалища. Эстрогенный гормон образуется в яичнике в течение всего цикла, но синтез его значительно возрастает в период развития фолликула. Под влиянием этого гормона (фолликулин) в матке усиливается синтез сократительного мышечного белка, увеличивается количество фосфорных соединений и гликогена, являющихся источником энергии, происходят сдвиги в обмене электролитов. |


