Развитие суставов. Возрастные изменения суставов Функции и задачи
Формирование суставов в антенатальном периоде, основные этапы развития которых отражены на рис. 5, связано с закладкой и дифференцировкой скелета.
На участке между двумя костями, где должен образоваться сустав, вначале имеется лишь неясно ограниченное предхрящевое скопление мезенхимы. Постепенно мезенхима становится бо-
Рис. 5. Развитие суставов:
1 – скопление мезенхимных клеток (предхрящевое состояние);
2 – место полости будущего сустава; 3 – надхрящница; 4 – надкостница;
5 – полость сустава; 6 – суставная капсула; 7 – суставной хрящ
лее плотной в местах, где должно начаться формирование хряща (I). Как только хрящевые модели будущих костей приобретают свойственную им форму, сустав намечается в виде расположенного между ними участка с меньшей концентрацией мезенхимы.
Надхрящница при своем формировании распространяется вокруг концов костей таким образом, что в месте образования сустава в течение некоторого времени имеется лишь рыхлая волокнистая соединительная ткань (II).
На третьем этапе в диафизах костей начинается процесс окостенения. Эпифизы еще остаются хрящевыми (III).
Разрыхление и, наконец, исчезновение соединительной ткани, расположенной между эпифизами, создает полость сустава в виде суставной щели. В хрящевых эпифизах появляются центры окостенения (IV).
Суставная капсула и связки сустава начинают формироваться из прилегающей к будущему суставу мезенхимы очень рано еще до появления суставной щели. Процесс завершения их образования (дифференцировка синовиальной оболочки, укрепление капсулы более или менее толстыми пучками коллагеновых волокон и т.д.) заканчивается несколько позднее (V).
В некоторых суставах (коленный, височно-нижнечелюстной и др.) прослойка мезенхимы, делящая полость сустава на две суставные щели, замещается хрящевой тканью и превращается в суставной диск или мениск. Хрящевая губа, в отдельных суставах (плечевой, тазобедренный), формируется из внутрисуставного хряща, у которого резорбируется его центральная часть, а периферические отделы плотно прирастают к краю суставной поверхности кости.
Образование непрерывных соединений костей (синартрозов) происходит совершенно иначе. В мезенхимальной закладке не появляется полость, а, наоборот, соединительная ткань удерживает обе кости более или менее плотно. Такое соединение называется синдесмозом. Кости могут быть связаны друг с другом и хрящевой тканью, замещающей соединительную ткань – синхондроз. Если же соединительная или хрящевая ткань заменяется костью, то мы говорим о синостозе.
При образовании переходных форм соединений (полусуставы – симфизы) из мезенхимальной соединительно-тканной прослойки, между сочленяющимися костями, формируется хрящевая прослойка значительной толщины. В толще этого хряща, на месте среднего слоя, появляется небольших размеров щель.
ОБЩАЯ АРТРОЛОГИЯ
Непрерывные соединения
Прерывистые соединения
Классификация суставов
Биомеханика суставов
Артрология – учение о соединениях костей друг с другом. Вместе с остеологией и мио-
логией артрология составляет раздел анатомии об опорно-двигательном аппарате.
Артрология подразделяется на общую и частную. Первая изучает различные типы со-
единений костей, классифицирует отдельные части суставов, вторая описывает частные
формы соединений и сочленений между отдельными костями.
Все соединения делятся на две группы: непрерывные и прерывистые.
Непрерывные соединения
Непрерывные соединения, или синартрозы, являются филогенетически более древни-
ми и более просто устроены. В зависимости от вида ткани, которая участвует в соеди-
нении костей, их подразделяют на фиброзные, хрящевые и костные.
Фиброзные соединения , articulationes fibrosae , построены из оформленной плотной
волокнистой (фиброзной) соединительной ткани. Эти соединения, в свою очередь,
подразделяются на синдесмозы и швы.
К синдесмозам относят связки и мембраны (Рис. 1). Те и другие состоят из волнообраз-
но извитых пучков коллагеновых волокон и небольшого количества эластических воло-
кон. Коллагеновые волокна являются основными носителями механических свойств
связок и мембран. Они устроены аналогично волокнам костной ткани, имеют толщину
от 20 до 250 мкм и состоят из агрегатов молекул коллагена (мицеллярных цепочек).
Коллагеновые волокна обладают большой упругостью и слабой растяжимостью. Мо-
дуль упругости коллагеновых волокон составляет 10 000 кг/см2, то есть на один поря-
док ниже, чем у компактной кости. При растяжении коллагеновые волокна удлиняются
на 10-20% своей первоначальной длины. Предельная прочность на растяжение у кол-
лагена весьма велика и достигает 500-1000 кг/см2. Поэтому связки могут выдерживать
большую нагрузку. Например, подвздошно-бедренная связка выдерживает нагрузку до
Некоторые связки состоят из соединительной ткани, в которой преобладают эластиче-
ские волокна. Такого рода соединения известный анатом А.Раубер назвал в свое время
синэластозами. Эластические волокна обладают небольшой упругостью и большой
растяжимостью. Модуль упругости эластина составляет всего 6 кг/см2, поэтому эласти-
ческие волокна растягиваются при небольшой нагрузке. Они могут удлиняться в 2.5
раза и после снятия нагрузки возвращаются в исходное состояние. Благодаря своей
растяжимости эластические связки выполняют в опорно-двигательном аппарате рес-
сорную функцию. Например, желтые связки, соединяющие дуги позвонков. Они создают сильное эластическое напряжение на дорсальной стороне позвоночного столба и
способствуют его выпрямлению.
Швы представляют собой тонкие пластинки волокнистой соединительной ткани, рас-
положенные между краями костей черепа. В зависимости от характера краев костей
швы подразделяются на зубчатые (Рис. 2), чешуйчатые (Рис. 3) и плоские (Рис. 4).
Особым видом фиброзного соединения является вколачивание, гомфоз, gomphosis ,
или зубоальвеолярное соединение; корни зубов прикрепляются к зубной альвеоле по-
средством соединительнотканных волокон (Рис. 5).
Кроме того, к фиброзным соединениям относят роднички черепа, которые имеются
только у плодов и новорожденных.
Хрящевые соединения , articulationes cartilagineae , называют также синхондрозами
(Рис. 6). В соединениях костей встречается два вида хряща: гиалиновый и волокнистый,
или коллагеновый. Подобно тому как кость покрыта надкостницей, поверхность хряща
покрыта надхрящницей, перихондрием. Расположенные в перихондрии соединитель-
нотканные волокна имеют форму аркад и продолжаются в волокна самого хряща.
Хрящевая ткань характеризуется значительной упругостью и растяжимостью. Межпо-
звоночные диски при сжатии выдерживают 800-2200 кг у мужчин и 500-1000 кг у жен-
щин; предельная нагрузка при растяжении составляет 150-225 кг. При этом диски уд-
линяются на 50-60% своей первоначальной длины.
К хрящевым соединениям относятся синхондрозы черепа, расположенные между кос-
тями основания черепа, а также синхондрозы грудины, соединяющие с телом грудины
ее рукоятку и мечевидный отросток (Рис. 7). Большинство синхондрозов являются вре-
менными. Они существуют только до определенного возраста, а затем хрящевая ткань
заменяется костной, то есть образуется синостоз (Рис. 8).
В качестве особого вида хрящевого соединения выделяют симфиз , symphysis (Рис. 9).
Он отличается от синхондроза тем, что в хряще имеется небольшая щелевидная по-
лость. Симфиз представляет как бы переходную форму от непрерывных соединений к
прерывистым. К симфизам относят сращение лобковых костей между собой. Полагают,
что образование полости в хряще происходит вследствие его растяжения. В межпозво-
ночных дисках также находятся небольшие полости, поэтому в настоящее время их на-
зывают межпозвоночными симфизами (Рис. 10).
Прерывистые соединения
Прерывистые соединения костей, или диартрозы, отличаются не только большей
сложностью строения, но и функциональными качествами. В противоположность ма-
лоподвижным или совсем неподвижным непрерывным соединениям диартрозы до-
пускают многообразные и направленные движения звеньев скелета. Возможность
дифференцированных движений головы и конечностей у наземных позвоночных оп-
ределяется степенью развития прерывистых соединений в их скелете.
К диартрозам относятся синовиальные соединения, articulationes synoviales , обычно
называемые суставами, articulatio . Сустав представляет собой орган, в построении ко-
торого принимают участие хрящевая, костная и собственно соединительная ткань. В
строении сустава можно выделить основные и вспомогательные элементы.
К основным элементам, которые имеются в любом синовиальном соединении, следует
отнести суставные поверхности, суставной хрящ, суставную полость, суставную капсулу
и синовиальную жидкость (Рис. 11).
Суставные поверхности , facies articulares , располагаются на костях, участвующих в об-
разовании сустава. Каждый сустав содержит, по крайней мере, одну пару сочленяю-
щихся поверхностей. Одна из них, как правило, выпуклая, это – суставная головка, а
другая – вогнутая – суставная впадина. Выпуклая поверхность имеет всегда большую
протяженность, чем вогнутая.
Суставные поверхности покрывает суставной хрящ , cartilago articularis . Толщина хряща
варьирует от 0.2 до 1.5 мм. На суставной впадине хрящ мягче, чем на суставной голов-
ке. Большинство суставных поверхностей покрыты гиалиновым хрящом, и лишь в неко-
торых суставах, например височно-нижнечелюстном и грудино-ключичном, имеется
волокнистый хрящ. По краю суставного хряща фиброзный слой надкостницы продол-
жается непосредственно в поверхностный слой волокон самого хряща. Таким образом,
вся кость вместе с суставным хрящом окружена единой фиброзной оболочкой. Наруж-
ная поверхность хряща гладкая, и это позволяет суставным поверхностям легко пере-
мещаться относительно друг друга. Благодаря своей эластичности суставной хрящ пре-
дохраняет концы костей от повреждения при толчках и сотрясениях.
Суставная капсула , capsula articularis , охватывает части костей, которые принадлежат
суставу. Капсула прикрепляется по краям суставных поверхностей или несколько отсту-
пя от них и герметично закрывает сустав. Суставная капсула состоит из двух мембран:
фиброзной и синовиальной. Фиброзная мембрана образует наружный слой. Она со-
стоит из волокнистой соединительной ткани, содержащей много коллагеновых воло-
кон. В фиброзную мембрану вплетаются связки, укрепляющие сустав; в этих местах сус-
тавная капсула бывает утолщена. В местах, свободный от прикрепления связок, фиб-
розная мембрана более тонкая, и здесь могут образовываться выпячивания суставной
капсулы. Синовиальная мембрана представляет внутренний слой суставной капсулы.
Она покрывает все образования, находящиеся в суставе, за исключением суставных
хрящей. Эта оболочка тонкая, рыхло соединена с фиброзной и потому подвижна, со-
держит коллагеновые и эластические волокна. В некоторых суставах синовиальная
мембрана образует складки, plicae synoviales , которые содержат жировую ткань и вда-
ются в полость сустава, заполняя в ней свободные участки. Более мелкие выросты – си-
новиальные ворсинки, villi synoviales , увеличивают поверхность синовиальной мембра-
ны, что, по-видимому, имеет значение для обменных процессов в суставе. Синовиаль-
ная мембрана богато снабжена кровеносными и лимфатическими сосудами и нерва-
Синовиальная мембрана и суставные поверхности ограничивают суставную полость ,
cavitas articularis , которая обычных условиях имеет вид узкой щели, и лишь при забо-
леваниях, когда в суставе скапливается большое количество жидкости, объем сустав-
ной полости увеличивается настолько, что суставные поверхности могут разойтись.
Синовиальная жидкость , или синовия, synovia , вырабатывается синовиальной мем-
браной. В норме она содержится в полости сустава в небольшом количестве. Синовия
играет роль смазки в суставах. Трение в суставах весьма значительно, коэффициент
трения равен 0.01, то есть имеет такую же величину, как в смазываемых подшипниках.
Однако способ смазки в суставах иной, чем те, которые применяются в технике. Как по-
казали исследования, суставной хрящ напоминает губку, с очень тонкими порами и
пропитан синовией, которая может быть из него выжата. Такая структура суставного
хряща позволяет осуществить неизвестную в технике «выжимающую смазку». Чтобы
выжимающая смазка была достаточно эффективной, необходимо постоянное неболь-
шое перемещение суставных поверхностей. Замечательными свойствами синовии яв-
ляются ее вязкость и упругость. Вязкость синовиальной жидкости зависит от одного из
ее компонентов – гиалуроновой кислоты, которая входит в состав основного вещества
соединительной ткани. При движениях в суставе вязкость синовии уменьшается в пять-
сот раз; благодаря этому облегчается работа сустава. Упругость синовии можно проде-
монстрировать с помощью простого опыта. Если нанести на стеклянную пластинку кап-
лю жидкости, взятой из сустава, и наложить сверху выпуклую линзу, то сближение лин-
зы с пластинкой прекратится, когда между ними еще остается некоторый промежуток.
Тонкий слой синовии между двумя поверхностями ведет себя как пленка из тонкой ре-
зины. Таким образом обеспечивается постоянной разделение суставных поверхностей,
что играет важную роль в механизме сустава. Синовия выполняет также защитную и
метаболическую функции, обеспечивает трофику суставного хряща, который лишен со-
судов. Синовиальная жидкость участвует в обмене веществ между содержимым суста-
ва и сосудистым руслом синовиальной мембраны. При введении в полость сустава
различных веществ они всасываются в сосуды синовиальной мембраны, причем дви-
жения в суставе значительно ускоряют всасывание. Между синовиальной мембраной,
синовиальной жидкостью и суставным хрящом существует морфологическая и функ-
циональная общность. На этом основании объединяют эти компоненты и выделяют
понятие «синовиальная среда сустава».
К вспомогательным элементам сустава относят суставные диски, мениски, губы, связки,
синовиальные сумки.
Суставной диск , discus articularis , представляет собой пластинку из волокнистого хря-
ща, покрытую синовиальной мембраной, которая располагается в полости сустава ме-
жду поверхностями сочленяющихся костей и срастается с суставной капсулой. Подоб-
ные образования имеются в височно-нижнечелюстном, грудино-ключичном и лучеза-
пястном суставах (Рис. 12).
Разновидностью диска являются суставные мениски , menisci articulares , находящиеся в
коленном суставе. Они представляют собой изогнутые хрящевые пластинки полулун-
ной и серповидной формы, укрепленные в суставе с помощью особых связок (Рис. 13).
Суставные диски и мениски ввиду своей эластичности смягчают удары и сотрясения,
передающиеся на сустав. Они также играют определенную роль в механизме движе-
Суставная губа , labrum articulare , представляет собой кольцевидное образование из
волокнистого хряща, которое прикрепляется по краю суставной впадины, углубляя ее и
увеличивая ее поверхность. Суставные губы имеются в плечевом, тазобедренном и не-
которых других суставах (Рис. 14).
Большую роль в укреплении суставов и движениях в них играют связки . Совокупность
их образует связочный аппарат сустава (Рис. 15). По отношению к суставной капсуле
выделяют 3 вида связок:
1. Внекапсульные связки, ligamenta extracapsularia – располагаются вне суставной
капсулы, но часто вплетаются в нее.
2. Капсульные связки, ligamenta capsularia – представляют собой утолщения сустав-
ной капсулы.
3. Внутрикапсульные связки, ligamenta intracapsularia – находятся в суставной полос-
ти и покрыты синовиальной мембраной (Рис. 16).
Вместе с суставной капсулой и мышцами связки обеспечивают укрепление суставов и
контакт суставных поверхностей костей. Многие связки тормозят и ограничивают дви-
жения в суставах. Имеются направляющие связки, которые оказывают влияние на ход
движения в суставе, взаимодействуя при этом с другими его частями, например локте-
вая коллатеральная связка локтевого сустава. У ряда связок указанные функции соче-
таются. Например медиальная (дельтовидная) связка голеностопного сустава выполня-
ет укрепляющую, тормозящую и направляющую функции.
Можно выделить несколько закономерностей расположения связок:
1. Связки распределяются в каждом суставе в зависимости от числа и положения его
осей вращения.
2. Связки располагаются перпендикулярно данной оси вращения и преимущественно
по ее концам.
3. Связки лежат в плоскости данного движения сустава.
Синовиальные сумки , bursae synoviales , представляют собой выпячивания синовиаль-
ной мембраны в истонченных участках фиброзной оболочки сустава. Размеры и форма
синовиальных сумок различны (Рис. 17). Как правило, синовиальные сумки располага-
ются между поверхностью кости и движущимися возле нее сухожилиями мышц. Сумки
устраняют трение друг о друга соприкасающихся поверхностей сухожилий, костей.
Таковы общие черты строения суставов. Одним из основных моментов, определяющих
целостность сустава, является постоянный контакт суставных поверхностей как в покое,
так и в движении.
Объем движений в каждом суставе зависит от целого ряда факторов.
1. Разность площадей сочленяющихся суставных поверхностей – главный фактор.
Из всех суставов наибольшая разность площадей суставных поверхностей в плече-
вом суставе (площадь головки плечевой кости в 6 раз больше площади суставной
впадины на лопатке), поэтому в плечевом суставе самый большой объем движе-
ний. В крестцово-подвздошном сочленении суставные поверхности по площади
равны, поэтому движения в нем практически отсутствуют.
2. Наличие вспомогательных элементов . Например, мениски и диски, увеличивая
конгруэнтность суставных поверхностей, увеличивают объем движений. Суставные
губы, увеличивая площадь суставной поверхности, способствуют ограничению
движений. Внутрисуставные связки ограничивают движения только в определен-
ном направлении (крестообразные связки коленного сустава не препятствуют сги-
банию, но противодействуют чрезмерному разгибанию).
3. Комбинация суставов . У комбинированных суставов движения определяются по
суставу, имеющему меньшее число осей вращения. Хотя многие суставы, исходя из
формы суставных поверхностей, способны выполнять больший объем движений,
но он у них ограничен из-за комбинации. Например, по форме суставных поверхностей латеральные атлантоосевые суставы – плоские, но в результате комбина-
ции со срединным атлантоосевым суставом они работают как вращательные. Это
же относится и к суставам ребер, суставу кисти, суставу стопы и др.
4. Состояние капсулы сустава . При тонкой, эластичной капсуле движения соверша-
ются в большем объеме. Даже неравномерная толщина капсулы в одном и том же
суставе сказывается на его работе. Например, в височно-нижнечелюстном суставе
капсула тоньше спереди, чем сзади и сбоку, поэтому наибольшая подвижность в
нем именно кпереди.
5. Укрепление капсулы сустава связками . Связки оказывают тормозящее и направ-
ляющее действие, так как коллагеновые волокна обладают не только большой
прочностью, но и малой растяжимостью. В тазобедренном суставе подвздошно-
бедренная связка препятствует разгибанию и повороту конечности кнутри, лобко-
во-бедренная связка – отведению и вращению наружу. Самые мощные связки на-
ходятся в крестцово-подвздошном суставе, поэтому движений в нем практически
6. Мышцы, окружающие сустав . Обладая постоянным тонусом, они скрепляют,
сближают и фиксируют сочленяющиеся кости. Сила мышечной тяги составляет до
10 кг на 1 см поперечника мышцы. Если удалить мышцы, оставить связки и капсу-
лу, то объем движений резко возрастает. Кроме непосредственного тормозящего
действия на движения в суставах, мышцы оказывают и косвенное – через связки,
от которых они начинаются. Мышцы при своем сокращении делают связки непо-
датливыми, упругими. Однако электромиографические исследования показали,
что стабильность суставов в определенных положениях достигается за счет связоч-
ного аппарата при минимальной активности мышц.
7. Синовиальная жидкость . Она оказывает сцепляющее воздействие и смазывает
суставные поверхности. При артрозоартритах, когда нарушается выделение сино-
виальной жидкости, в суставах появляются боль, хруст, объем движений уменьша-
8. Винтовое отклонение . Имеется только в плечелоктевом суставе и оказывает тор-
мозящее воздействие при движениях.
9. Атмосферное давление . Благодаря герметичности суставов в суставной полости
поддерживается отрицательное давление, равное 60-120 мм водяного столба. Ат-
мосферное давление способствует соприкосновению суставных поверхностей с
силой 1 кг на 1 см2, оказывает равномерное стягивающее воздействие, следова-
тельно, умеренно ограничивает движения. Например, в тазобедренном суставе эта
сила достигает 25 кг. Если изолировать сустав, удалить все мышцы и связки, то со-
единение суставных поверхностей сохраняется. Чтобы суставные поверхности ра-
зошлись, нужно рассечь суставную капсулу или ввести под давлением газ в по-
лость сустава.
10. Состояние кожи и подкожной жировой клетчатки . У тучных людей объем движе-
ний всегда меньше из-за обильной подкожной жировой клетчатки. У стройных,
подтянутых людей, у спортсменов движения совершаются в большем объеме. При
заболеваниях кожи, когда теряется эластичность, движения резко уменьшаются, а
нередко после тяжелых ожогов, ранений образуются контрактуры, значительно
препятствующие движениям.
Классификация суставов
Классификация суставов основывается на анатомических и функциональных признаках.
В зависимости от числа сочленяющихся поверхностей выделяют суставы простые и
Простой сустав , articulatio simplex , имеет только одну пару суставных поверхностей.
Большая часть суставов человека относится к простым, например межфаланговые
Сложный сустав , articulatio composita , включает две и более пар суставных поверхно-
стей, например локтевой (Рис. 19).
По функциональному признаку выделяют комплексные и комбинированные суставы.
Комплексным суставом, articulatio complexa , называют такой сустав, полость которого
полностью или частично разделена на две части суставным диском или мениском. На-
пример височно-нижнечелюстной сустав (Рис. 12).
Комбинированные суставы , articulationes combinatae – это анатомически изолирован-
ные суставы, которые всегда вместе участвуют в движениях. Например правый и левый
височно-нижнечелюстные суставы.
Рассматривая суставы, можно видеть, что суставные поверхности имеют различную
форму. Принято сравнивать форму суставных поверхностей с известными геометриче-
скими телами – шаром, эллипсоидом, цилиндром. Однако идеальных геометрических
поверхностей в организме не существует, поэтому уподобление суставов геометриче-
ским телам в какой-то мере условно. Все суставные поверхности обладают некоторой
кривизной, совершенно плоских поверхностей нет.
По форме суставных поверхностей выделяют следующие виды суставов:
1. Плоский сустав , articulatio plana – его суставные поверхности можно рассматри-
вать как участки шара большого радиуса. К плоским суставам относят дугоотрост-
чатые, запястно-пястные суставы II – V пальцев, крестцово-подвздошный, межбер-
цовый, предплюсне-плюсневые суставы (Рис. 20).
2. Шаровидный сустав , articulatio spheroidea – также не совсем отвечает своему на-
званию, так как его поверхности в разных участках имеют неодинаковую кривизну.
Шаровидными являются сустав головки ребра, плечевой, плечелучевой, таранно-
ладьевидный суставы (Рис. 21).
3. Чашеобразный сустав , articulatio cotylica – представляет собой разновидность ша-
ровидного. Имеется только один такой сустав – тазобедренный (Рис. 22).
4. Эллипсоидный сустав , articulatio ellipsoidea – поверхности его можно сравнить с
куском яичной скорлупы. К данному виду относятся ключично-акромиальный, лу-
чезапястный суставы. Эллипсоидные поверхности, как и сфероидные, или вогнуты,
или выпуклы во всех направлениях (Рис. 23).
5. Седловидный сустав , articulatio sellaris – обладает противоположным свойством,
суставные поверхности в одном направлении выпуклы, а в противоположном – вогнуты. Седловидными являются грудино-ключичный, запястно-пястный сустав I
пальца, пяточно-кубовидный сустав (Рис. 24).
6. Мыщелковый сустав , articulatio condylaris – соединение, при котором одна кость
сочленяется с другой посредством двух раздельных поверхностей. Каждая из этих
суставных поверхностей носит название мыщелка, независимо от того, является
она выпуклой или вогнутой. К мыщелковым суставам относят коленный (Рис. 25).
7. Блоковидный сустав , ginglymus – сустав с цилиндрическими суставными поверх-
ностями. Ось выпуклой суставной поверхности перпендикулярна оси самой кости,
а блок имеет небольшой гребешок, который направляет его движение. Таковыми
являются межфаланговые суставы, плечелоктевой и голеностопный (Рис. 26).
8. Цилиндрический сустав , articulatio trochoidea – ось выпуклой суставной поверхно-
сти идет в направлении продольной оси самой кости, а не перпендикулярно ей,
как в блоковидном суставе. К вращательным относят атлантоосевой, реберно-
поперечный, лучелоктевые и подтаранный суставы (Рис. 27).
Биомеханика суставов
Форма суставов стоит в тесной связи с их функцией. В учении о суставах находит свое
наглядное выражение диалектическое положение о единстве и взаимообусловленно-
сти строения и функции. Изучение движений в суставах – артрокинематика – является
одним из разделов биомеханики.
В суставах осуществляется движение костей относительно друг друга. Каждая отдельно
взятая кость, если рассматривать ее как физическое тело, может совершать поступа-
тельные движения по трем направлениям и вращаться вокруг трех взаимно перпенди-
кулярных осей. Соответственно этому она имеет 6 степеней свободы. В скелете кость
утрачивает часть степеней свободы, поскольку суставы позволяют осуществлять лишь
вращательные движения вокруг одной, двух или трех осей. Количеством осей враще-
ния и определяется число степеней свободы отдельных звеньев скелета.
В анатомии выделяют сагиттальную, фронтальную и вертикальную оси. Движения,
осуществляемые вокруг названных осей, определяют как сгибание (flexio ) и разгибание
(extensio ) вокруг фронтальной оси, отведение (abductio ) и приведение (adductio ) вокруг
сагиттальной оси, и собственно вращение (rotatio ) вокруг вертикальной оси. В качестве
особого вида рассматривают круговое движение (circumductio ), при котором перифе-
рический конец кости движется по окружности.
При любом движении, кроме вращения вокруг собственной оси, каждая точка кости
описывает в пространстве некоторую кривую линию. Если взять точку, находящуюся на
механической оси кости, то все ее движения совершаются в определенной плоскости,
которая всегда выпукла со стороны, противоположной суставу. Эта плоскость пред-
ставляет собой сферу, или овоид, движения. Протяженность овоида движения зависит
от амплитуды движений в суставе. Это понятие помогает описывать и графически
представлять движения в суставах (Рис. 28, Рис. 29, Рис. 30).
Число степеней свободы и типы движений в суставах зависят от формы суставных по-
верхностей. К одноосным суставам с одной степенью свободы относят блоковидные и
цилиндрические суставы. Двухосными с двумя степенями свободы являются эллипсоидные, седловидные и мыщелковые суставы. К многоосным суставам с тремя степенями свободы принадлежат шаровидные, чашеобразные и плоские суставы.
В некоторых руководствах, преимущественно старых, выделяется еще один вид суста-
вов – тугие суставы, или амфиартрозы. Под этим названием выделяется группа сочле-
нений с различной формой суставных поверхностей, но сходных по другим признакам:
они имеют короткую, туго натянутую суставную капсулу и очень крепкий, нерастяги-
вающийся вспомогательный аппарат. Тугие суставы смягчают толчки и сотрясения ме-
жду костями. В амфиартрозах движения имеют скользящий характер и крайне незна-
чительный объем.
Важное значение в артрокинематике имеет понятие о конгруэнтности суставов. Сустав-
ные поверхности имеют почти всегда различную площадь и кривизну. Поверхности, ко-
торые полностью соответствуют друг другу, называются конгруэнтными. Если такое со-
ответствие отсутствует, говорят о неконгруэнтных поверхностях. Если сравнить тазо-
бедренный и плечевой суставы, то можно видеть, что в тазобедренном суставе сочле-
няющиеся поверхности более подходят одна к другой, чем в плечевом суставе. Поэто-
му тазобедренный сустав является в большей степени конгруэнтным. Конгруэнтность
поверхностей в каждом суставе не является постоянной, она изменяется при движени-
ях и в зависимости от нагрузки. При нагружении сустава площадь контакта суставных
поверхностей возрастает, и конгруэнтность увеличивается. Это способствует более рав-
номерной передаче нагрузки на суставные концы костей.
Исходя из анализа конгруэнтности суставных поверхностей, Мак-Конейл различает в
каждом суставе замкнутое и разомкнутое положения (Рис. 32). Замкнутым является та-
кое положение, при котором достигается максимальная конгруэнтность сустава. При
замкнутом положении связки, укрепляющие сустав, натянуты и напряжены, они при-
жимают суставные поверхности друг к другу и полностью проявляют свою стабилизи-
рующую функцию. Сустав в замкнутом положении максимально устойчив, количество
степеней свободы в нем падает до нуля. При всех других положениях сустав является
разомкнутым. При этом суставные поверхности становятся неконгруэнтными, связки
расслабляются и могут быть реализованы все степени свободы для данного сустава.
Рассмотрим эти положения применительно к конкретным суставам. В плечевом суста-
ве замкнутое положение достигается при отведении и вращении наружу плечевой кос-
ти. При этом плечевая кость стабилизируется и может перемещаться только вместе с
лопаткой. Замыкание локтевого сустава происходит при разгибании и супинации. В лу-
чезапястном суставе замкнутое положение соответствует полному разгибанию кисти;
подвижность в суставе при этом отсутствует. У коленного и голеностопного суставов
замкнутым также является положение полного разгибания. Чтобы восстановить под-
вижность в суставе, его нужно привести в разомкнутое положение. Во всех приведен-
ных случаях это достигается сгибанием в сочетании с небольшим вращением внутрь.
При замыкании суставов создаются условия, способствующие переломам костей при
травмах, ввиду того, что не может проявиться рессорное действие соединений. Так,
перелом лучевой кости чаще всего происходит при падении на вытянутую руку с разо-
гнутой кистью, когда лучезапястный и локтевой суставы находятся в замкнутом поло-
Описываемые в учебниках виды движений в суставах редко осуществляются в своей
элементарной форме. Большинство движений являются сложными. Даже в таком, ка-
залась бы, простом случае, как движения ногтевых фаланг пальцев, можно заметить,
что при сгибании они слегка супинированы, а разгибание сопровождается пронацией
фаланг. Сочетание сгибания и разгибания с некоторой степенью вращения характерно
и для других блоковидных суставов. Например, в локтевом суставе при полном разги-
бании происходит пронация локтевой кости, а при сгибании она супинируется. Благо-
даря комбинации сгибания и разгибания с вращением блоковидный сустав выводится
из замкнутого положения и снова приводится в такое положение. Подобного рода за-
мыкающие и размыкающие движения относятся к обычным движениям в суставах.
Распространенным видом сложных движений является последовательное движение.
При этом часть тела, например конечность, последовательно переводится из одного
положения в другое и в результате серии движений может вернуться в исходное со-
стояние. В данном случае говорят об эргономическом цикле. Подобные циклы харак-
терны для различного рода повторяющихся рабочих движений.
Различают два вида вращательных движений: сочетанные и независимые. Сочетанное
вращение имеет место при осуществлении последовательных движений. Чтобы вы-
явить сочетанное вращение, нужно опустить руку с полупронированным предплечьем
так, чтобы ладонь была обращена к бедру. Затем рука поднимается вперед до горизон-
тального уровня и отводится в сторону на 90°, причем сохраняется полупронированное
положение предплечья. После этого рука приводится к туловищу, и оказывается, что
теперь она повернута к бедру уже не ладонью, а локтевым краем. Это значит, что в хо-
де последовательных движений произошло вращение наружу в плечевом суставе на
90°. Если из нового положения повторить тот же цикл движений, то снова произойдет
поворот руки на 90°, и кисть будет обращена к бедру своей тыльной стороной. Произ-
вести движения в третий раз, очевидно, уже не удастся. Таким образом было получено
вращение в результате серии движений, которые сами по себе не являются враща-
тельными. Такое сочетанное вращение возможно в любом суставе, имеющем 2 или 3
степени свободы. Всякое другое вращение называется независимым вращением.
Развитие и возрастные изменения суставов
Образование суставов в онтогенезе тесно связано с развитием костей. Из лекции по
общей остеологии известно, что скелет проходит бластемную, хрящевую и костную
стадии. При формировании хрящевых закладок костей в мезенхимальной бластеме ос-
таются промежуточные зоны, в которых не происходит образования хряща. В этих мес-
тах и развиваются суставы (Рис. 33). Зоны формирования плечевого и локтевого суста-
вов намечаются на 6-й неделе внутриутробного развития у эмбриона длиной 12 мм,
зоны тазобедренного и коленного суставов – у эмбриона длиной 13 мм. У эмбриона
длиной 14 мм уже вырисовываются характерные очертания суставных концов костей.
На 7-й неделе у эмбриона длиной 16-20 мм происходит дифференцировка основных
элементов сустава, отчетливо различаются промежуточная зона, суставная капсула с ее
фиброзной и синовиальной мембранами и перихондрий. Начинается формирование
суставного хряща. Путем разжижения центральной части промежуточной зоны образу-
ется суставная полость. В различных суставах появление полости происходит неодно-
временно. На 6-7-й неделе образуется полость в плечевом и коленном суставах, на 8-9-
й неделе – в локтевом и лучезапястном, на 10-11-й неделе – в височно-
нижнечелюстном и голеностопном и т.д. Внутрисуставные образования (диски, мени-
ски) закладываются на месте, а не мигрируют в полость сустава извне.
У новорожденного ребенка все элементы сустава анатомически сформированы, одна-
ко их тканевая структура значительно отличается от окончательной. Суставные концы
костей при рождении целиком состоят из хряща, окостенение большинства эпифизов
начинается на 1-м или 2-м году жизни и продолжается до пубертатного периода. Сус-
тавной хрящ у новорожденных имеет волокнистое строение. Перестройка хряща идет
очень интенсивно в первые три года жизни, а затем она замедляется и окончательно
затухает в период с 9 до 14 лет. К 14-16 годам суставной хрящ приобретает строение
типичного гиалинового. В синовиальной мембране после рождения увеличиваются
число и размеры складок и ворсин, происходит развитие сосудистой сети и нервных
окончаний. В возрасте 6-10 лет усложняется строение ворсинок, часть их приобретает
разветвленную форму. С 3 до 8 лет наблюдается усиленная коллагенизация суставной
капсулы и связок. В подростковом возрасте происходит утолщение суставной капсулы.
В 15-16 лет все внутрисуставные образования становятся хрящевыми. Окончательного
развития суставы, как и кости, достигают к 22-25 годам.
В пожилом и старческом возрасте в суставно-связочном аппарате происходят значи-
тельные изменения, в основе которых лежат глубокие ультраструктурные и биохими-
ческие процессы, протекающие в соединительной ткани. Они заключаются в обедне-
нии тканей водой, уменьшении содержания клеток и нарастании количества волокни-
стых структур, изменении свойств коллагена, дегенерации эластических волокон. Ос-
новное вещество хряща начинает изменяться уже в третьем десятилетии жизни. В по-
жилом возрасте идет процесс обызвествления суставных хрящей, а в старческом – в
них может происходить отложение кости. Суставные хрящи становятся тоньше. Изме-
нения в суставном хряще, капсуле и связках приводит к уменьшению объема движений
в суставах. Аналогичные изменения описаны в межпозвоночных дисках. Сопротивле-
ние дисков сжатию значительно снижается после 60 лет. Снижается также предел
прочности на разрыв связок, укрепляющих крупные суставы.
Аномалии суставов связаны с нарушениями развития костей и мышц. Недоразвитие
суставной впадины или головки приводит к врожденному вывиху, например в плече-
Формирование суставов в онтогенезе человека связано со сроками окостенения участвующих в их образовании костей, определяющих форму и рельеф суставных поверхностей, а также с особенностями развития суставных капсул и связок.
К моменту рождения все суставы имеют основные анатомические структуры, подвергающиеся в течение жизни изменениям. Ведущая роль в процессе их дальнейшего формирования и развития принадлежит силе тяжести вышележащих частей тела (статическим нагрузкам) и уровню физической активности в различных возрастных периодах.
Грудино-ключичный сустав . Грудинный конец ключицы у новорожденного имеет шаровидную форму; седловидный изгиб его начинает формироваться на первом году жизни, к 2-3 годам он хорошо выражен, а к 5-7 годам имеет форму такую же, как у взрослого, в дальнейшем увеличиваясь в размерах. Суставная поверхность рукоятки грудины растет, одновременно углубляясь, такими же темпами. Одновременно с суставными концами костей растет и диск между ними, но медленнее. Толщина диска с возрастом уменьшается, он плотнее сростается с капсулой сустава. Капсула постепнно утолщается. К 10-12 годам заметно утолщаются укрепляющие ее связки. Окончательного развития все элементы сустава достигают к 18-20 годам.
Акромиально-ключичный сустав . Его щелевидная полость у новорожденного относительно больше, чем у взрослого, что при слабости капсулы и связок его обеспечивает и больший, чем у взрослого, объем движений в суставе. Утолщение капсулы и связок начинается с 2-3 лет. К 12-14 годам сустав достигает дефинитивного состояния.
Плечевой сустав . У новорожденного он образован полностью хрящевой суставной впадиной лопатки, имеющей овальную форму, с плоской поверхностью и невысокой хрящевой губой, а также полностью хрящевой шаровидной головкой плечевой кости, имеющей с телом (диафизом) кости угол 170°(на 30° больше, чем у взрослого). Суставная капсула тесная, сильно натянута. В нижнем ее отделе имеются складки. Из-за неравномерного расположения волокнистых структур ее толщина в разных отделах неодинакова. Клювовидно-плечевая связка, идущая от основания клювовидного отростка лопатки и вплетающаяся в капсулу сустава, короткая, плотная, хорошо развита. Клювовидно-акромиальная связка лопатки имеет косое направление и тесно прилежит к капсуле сустава. Все эти анатомические особенности: положение отростков лопатки, связок, угол наклона эпифиза плеча к его диафизу, малые размеры капсулы резко ограничивают отведение плеча, вращение его вокруг вертикальной оси.
Развитие костей и нарастающая двигательная активность изменяют структуру плечевого сустава. К 4-м годам суставная впадина лопатки увеличивается в размерах и приобретает овоидную форму и вогнутость, свойственную взрослому, разрастается хрящевая суставная губа. Полное формирование суставной впадины лопатки заканчивается к концу пубертатного периода - к 15-16 годам. К 10-12-летнему возрасту клювовидный отросток лопатки занимает типичное положение, смещаясь вверх, что изменяет положение клювовидно-акромиальной связки. Угол между головкой и телом плечевой кости уменьшается до 140°. Это способствует увеличению объема движений. Максимум подвижности в суставе приходится на 8-10 лет. В укреплении плечевого сустава ведущая роль принадлежит мышцам. Будучи рыхло связанными с капсулой в первые месяцы жизни, они в дальнейшем своими сухожильными частями вплетаются в нее по всей поверхности, за исключением нижнего отдела, остающегося наиболее слабым местом капсулы.
Подвижность в соединениях пояса верхней конечности и в плечевом суставе увеличивается до 10-12 лет, до 16 лет показатели ее сохраняются на высоком уровне, а затем начинают снижаться, особенно резко после 50 лет.
Локтевой сустав . К моменту рождения концы плечевой, локтевой и лучевой костей хрящевые. Венечная ямка, локтевая и лучевая ямки плеча и борозда блока выражены слабо. Это же относится и к лучевой вырезке локтевой кости. Слабо развит локтевой отросток. Размеры суставной капсулы относительно велики, ее толщина спереди больше, чем сзади. Коллатеральные связки развиты слабо и еще не дифференцируются от суставной капсулы. Кольцевая связка лучевой кости слабая. Просторная капсула образует вдавление в полость сустава в области локтевой, лучевой и венечной ямок, а между плечевой и лучевой костями образуется своеобразный мениск - складка синовиальной оболочки с подлежащей жировой тканью. Самое слабое место капсулы - область ее прикрепления к краям венечной ямки.
На 1-м году жизни складка (мениск), разделявшая плечевую и лучевую кости, исчезает. В раннем детстве капсула остается просторной, коллатеральные связки удлиняются, объем движений возрастает. Анатомическая структура локтевого отростка в раннем детстве создает предпосылки для переразгибания предплечья в суставе. Сохраняющаяся тонкость и слабость кольцевой связки, а также скошенность головки лучевой кости являются условиями для выскальзывания последней из связки, в результате чего возможны подвывихи в плечелучевом суставе у детей раннего возраста. Большую роль в укреплении локтевого сустава в первые годы жизни играет мышца-супинатор. В ней два слоя - поверхностный и глубокий. Сухожильные волокна глубокого слоя вплетаются в капсулу сустава с лучевой стороны, укрепляя коллатеральную лучевую связку.
К 16-ти годам капсула и связочный аппарат сустава достигают дефинитивного состояния. Сгибательно-разгибательная подвижность увеличивается в локтевом суставе до 11-12 лет и долго сохраняется примерно на одном уровне, снижаясь после 40 лет. Пронаторно-супинаторная подвижность предплечья увеличивается только до 8-10 лет.
Ранние занятия спортом (гимнастикой) при слабости мышц, принимающих участие в укреплении локтевого сустава, и незавершенности его развития при больших нагрузках могут привести к деформации сустава, нежелательному переразгибанию, сохраняющемуся на всю жизнь, что создает дополнительную нагрузку на мышцы-сгибатели предплечья в локтевом суставе, способствует повреждению капсулы, вывихам, переломам костей.
Лучезапястный сустав . На первом году жизни он образован хрящевым дистальным эпифизом лучевой кости и хрящевыми телами развивающихся костей проксимального ряда запястья - ладьевидной, полулунной и трехгранной. Его суставная капсула тесно натянута в большей степени с ладонной стороны, что ограничивает разгибание кисти. Связки плохо дифференцированы от капсулы. Дальнейшее формирование сустава определяется сроками окостенения образующих его костей.
На первых годам практически отсутствует, становясь выраженным к 7-ми годам, а полностью формируясь к 10-13-ти годам, канал кисти - углубление на ладонной поверхности сустава, служащее ложем для сухожилий мышц-сгибателей. Канал кисти с его поперечной связкой создает достаточную опору идущим в нем сухожилиям к 16-ти годам.
Связочный аппарат сустава, в частности коллатеральные связки, достигают максимальной прочности к периоду полового созревания.
Капсула сустава в течение всей жизни имеет ряд слабых мест, преимущественно на тыльной стороне, что служит предпосылкой возникновения выпячиваний ее, при повреждении которых могут возникать воспалительные процессы.
Ранняя спортивная специализация без учета степени развития образующих сустав структур может существенно сказаться на его формировании. Так, при начале занятий гимнастикой с 5-6 лет типичная для сустава эллипсовидная форма может преобразоваться в шаровидную, обеспечивающую больший объем движений вокруг всех осей, включая вертикальную. В противоположность этому, у баскетболистов сустав становится менее подвижным, приобретая форму более вытянутого в поперечном направлении эллипса, что ограничивает боковые движения (вокруг сагиттальной оси), одновременно увеличивая объем сгибания и разгибания в общей сложности до 200°.
Увеличение активной подвижности в суставе наблюдается до 31-40 лет, объем пассивных движений кисти уменьшается после 8-9 лет.
Тазобедренный сустав . У новорожденного он образован преимущественно хрящевой вертлужной впадиной тазовой кости и полностью хрящевой головкой бедренной кости. Вертлужная впадина имеет овальную форму размером 18x14 мм, по краям ее расположена невысокая хрящевая губа. Сама впадина уплощена, особенно сглажен ее верхний край, что является предпосылкой для врожденных вывихов бедра. В связи с прямохождением и под действием сил тяжести объем вертлужной впадины быстро увеличивается. К 6-ти годам она приобретает округлые очертания (размер ее становится равным 31x30 мм), более высокой становится хрящевая губа. Быстро изменяется и бедренная кость. Укороченная поначалу ее шейка, расположенная при рождении под углом к телу кости в 150°, к 5-6-ти годам удлиняется, а величина угла уменьшается до 140°.
Капсула сустава у новорожденного тонкая, натянутая. Толщина ее переднего отдела – 1-2 мм, а сзади и снизу - 0,2-0,3 мм. Связки плохо дифференцируются. Синовиальная оболочка прикрепляется к наиболее выступающим частям суставной губы и по границе суставной поверхности головки бедра. Она менее обширная, чем фиброзная оболочка капсулы, прикрывающая также шейку бедренной кости и крепящаяся по межвертельному гребню и межвертельной линии. Синовиальная оболочка образует круговой карман вокруг вертлужной впадины, который покрывает край суставной губы, а на бедренной кости образует ряд складок в виде валика, содержащего значительное количество жировой ткани.
Связка головки бедра первично плоская, ее длина примерно равна диаметру головки бедра - 8-11 мм. В толще этой связки, а ширина ее у места начала в ямке вертлужной впадины – 4-7 мм и 4-6 мм – у головки бедра при толщине на всем протяжении равной 1-2 мм, проходит от 3 до 20 артерий небольшого калибра, питающих головку бедра. Связка выдерживает нагрузку при испытании прочности на разрыв до 7-8 кг.
Подвздошно-бедренная связка у новорожденного хорошо развита - толстая, треугольной формы, длиной около 1,5 см. Лобково-бедренная и седалищно-бедренная связки тонкие, едва намечены, плотно сращены с фиброзной оболочкой капсулы. В связи с особенностями расположения связок и строения впадины наиболее слабым местом сустава является верхнезадний отдел, что необходимо учитывать при воздействии избыточных вертикальных нагрузок в детском возрасте. Короткость и прочность подвздошно-бедренной связки при рождении резко ограничивает разгибание и вращение кнаружи бедра в тазобедренном суставе.
К концу 1-го года жизни с началом хождения подвздошно-бедренная связка удлиняется, капсула сустава становится несколько свободнее, размах движений в суставе увеличивается. Параллельно развиваются лобково-бедренная и седалищно-бедренная связки. Седалищно-бедренная связка, разрастаясь, покрывает капсулу сустава сзади и сверху к периоду раннего детства. В дальнейшем она продолжает утолщаться, достигая достаточной прочности к 10-12-ти годам.
Головка бедренной кости на 1-2-м году жизни постепенно погружается в суставную впадину, и круговая зона (внутрикапсульная связка) смещается с головки на шейку. Этот процесс заканчивается к 12-16-ти годам.
Тазобедренный сустав в основном принимает дефинитивное строение к 15-16 годам, однако, в течение всей жизни наиболее слабым местом его капсулы остается задненижний отдел, наименее укрепленный связками и мышцами, что редко, но при достаточно большом внешнем усилии при отведении в сочетании с вращением бедра внутрь (шпагат) у нетренированных людей может привести к вывиху.
В тазобедренном суставе наиболее интенсивный прирост подвижности характерен для младшего школьного возраста, в 12-15 лет ее изменения невелики, а с 16-ти лет подвижность сустава несколько снижается, стабилизируясь в интервале 20-50 лет и вновь уменьшаясь после 50 лет.
Ранняя специализация в различных видах спорта оказывает существенное влияние на формирование тазобедренного сустава. У футболистов размеры головки бедра и глубина вертлужной впадины, охватывающей головку, возрастают, что ограничивает объем движений в суставе. У гимнастов, наоборот, диаметр головки бедренной кости, меньше, а вертлужная впадина мельче. Объем суставной впадины оказывается значительно меньше головки, и размах движений в суставе большой.
Коленный сустав . Мениски сустава при рождении тонкие, эластичные и легко смещаются. Поперечная связка обычно хорошо выражена, а крестообразные - тонкие, короткие и почти одинаковой длины. Капсула сустава тонкая, особенно в верхнем отделе. Наружные связки слабые, особенно медиальная коллатеральная. Связка надколенника короткая. Синовиальные сумки малы или отсутствуют.
В период раннего детства изменяется положение проксимального эпифиза большой берцовой кости, и ее суставные поверхности занимают более горизонтальное положение (у новорожденного они обращены в большей степени назад). Неравномерно растущие мыщелки бедра из горизонтального положения переходят в косое. Наружный мыщелок постепенно опускается. Усиление этого процесса может привести к О-образному искривлению нижних конечностей, а в других случаях - к Х-образному, что особенно часто происходит при увеличении вертикальной нагрузки на коленные суставы в возрасте от 4-5-ти до 10-12-ти лет. Положение эпифизов бедра и большеберцовой кости устанавливается к 10-12-ти годам. К этому периоду завершается оссификация надколенника, дифференцирование связочного аппарата. Синовиальные сумки в окружности сустава развиваются по мере дифференцирования прикрепляющихся здесь мышц.
В коленном суставе сгибательно-разгибательная подвижность начинает снижаться уже с 7-ми лет. Пронаторно-супинаторная подвижность голени увеличивается до 10-11 лет, затем снижается.
Голеностопный сустав . У новорожденного он образован суставными поверхностями блока таранной кости, уже имеющей точки окостенения, и полностью хрящевых дистальных эпифизов большой берцовой и малой берцовой костей. Блок таранной кости имеет форму неправильной трапеции. Латеральная лодыжка отстоит кзади по сравнению с медиальной. Капсула сустава плотная. Связки хорошо выражены. Из них дельтовидная (медиальная) сравнительно более прочная, относительно короткая и затрудняет супинирование и подошвенное сгибание стопы.
Максимальной подвижности сустав достигает к 7-8-ми годам, когда практически завершается формирование его структур.
При систематических занятиях силовыми видами спорта с раннего возраста происходит уплощение блока таранной кости, значительно уменьшающее подвижность в суставе, что сказывается на устойчивости тела при выполнении характерных для этих видов спорта приемов и движений.
Стопа . Становление ее опорной и рессорной функции начинается, когда ребенок впервые самостоятельно принимает вертикальное положение. Рессорные свойства стопы определяются степенью выраженности ее сводов и их прочностью. Своды стопы, удерживаемые пассивными и активными затяжками, формируются еще до рождения ребенка. Стопа новорожденного выглядит плоской, так как на ней хорошо развит слой подкожной жировой клетчатки. Период активного формирования сводов приходится на возраст от 3 до 7 лет. В этот период наиболее часто возникает плоскостопие. В период второго детства (8-12 лет) скелет стопы фактически заканчивает формирование. Рост отдельных костей идет неравномерно. Прочность связочного аппарата значительно нарастает за счет изменения соотношения коллагеновых и эластических волокон в сторону увеличения первых. Интенсивный рост пяточной и всех плюсневых костей приводит к 12-летнему возрасту к качественно новому соотношению между отделами стопы, характерному для взрослого человека.
Окончательно рессорные свойства стопы, обусловленные степенью выраженности ее сводов, устанавливаются к 16-20 годам, после чего плоскостопие (если оно не появилось раньше), как правило, не развивается.
В возрастных изменениях подвижности стопы выделяют три этапа:
· от 1 года до 11-13 лет - уменьшение размаха движений;
· до 40 лет - стабилизация подвижности;
· после 40 лет - снижение подвижности, особенно прогрессирующее к 70 годам.
В последующее время основные закономерности органогенеза суставов были изучены достаточно подробно отечественными и зарубежными исследователями.
Результаты исследований, обобщенные в работах многих авторов, послужили основой для дальнейшего изучения органогенеза синовиальных суставов, который в настоящее время рассматривается как сложный многоэтапный процесс. В то же время клеточные и особенно молекулярные механизмы процесса становления синовиальных суставов остаются малоизученными.
В настоящее время идентифицирован набор регуляторных генов, которые, как полагают, контролируют организацию скелета развивающейся конечности. Выявлено также, что зона, где идет формирование сустава, оказывает организующее влияние на процесс развития скелета.
В последние годы большое внимание было уделено изучению роли костных морфогенетических белков (BMP) в развитии скелета у позвоночных. BMP относятся к большому семейству факторов роста и дифференцировки. Они определяют процесс становления скелета в целом и особенно формирование синовиальных суставов. Выявлено, что избыточная продукция BMP приводит к гиперпродукции ткани хряща и зарастанию полости формирующегося сустава. В постнатальном онтогенезе BMP сохраняют свое действие, обеспечивая нормальное функционирование сустава.
В условиях патологии функционирование BMP продолжается, но их действие изменено факторами, вызывающими воспаление (в частности, интерлейкинами).
Развитие парных конечностей у позвоночных: эмбриологический аспект
Процесс формирования конечности у человека и позвоночных животных можно описать следующим образом. После завершения процессов, связанных с гаструляцией, у зародышей человека и позвоночных животных наступает следующий этап развития, который называют нейруляцией, а саму стадию нейрулой. Этот период характеризуется тем, что в нем начинаются процессы построения отдельных систем органов, т.е. процессы органогенеза. Одним из таких органогенезов будет процесс развития парных закладок конечностей. Еще в процессе гаструляции в области формирования хорды обособляется клеточный материал, именуемый хордомезодермой. На следующем этапе обособляется мезодерма будущих сомитов в виде боковой пластинки.
У зародышей амфибий (анамнии) на ранних этапах развития зачатки конечностей представляют собой обособленные бугорки. У амниот зачатки конечностей образуются в виде длинных складок в теле зародыша, растянутых в переднезаднем направлении (вольфовы гребни). Средняя часть вольфовых гребней рассасывается, а из оставшихся передних и задних отделов их образуются передние и задние парные конечности. Вначале клетки эктодермального эпителия не принимают активного участия в формировании зачатка конечности. Закладка из клеток эктодермального эпителия пассивно растягивается быстро растущими клетками париетального листка мезодермы. Позже эктодермальная закладка начинает активно участвовать в росте конечности. На верхушке зачатка конечности эктодерма образует утолщение - апикальный гребешок. По мере роста зачатка конечности его форма меняется.
Еще в 1948 г. J.W. Saunders показал, что элементы скелета конечности закладываются и дифференцируются в проксимо-дистальном направлении. Этим же автором было установлено, что развитие конечности происходит благодаря взаимодействию между дистальной мезенхимой закладки конечности и эктодермой апикального гребешка. При этом клетки дистальной мезенхимы зачатка конечности, находясь в недифференцированном состоянии, формируют так называемую, активную зону, клетки которой обладают очень высокой способностью к пролиферации.
Одновременно с изменением внешней формы зачатка конечности идет формирование его внутреннего скелета. Первым формируется зачаток проксимального хряща - эпиплодия, из которого образуются хрящевые модели соответственно плечевой и бедренной костей. Затем возникает следующий зачаток - зигоплодий, из которого образуются хрящевые модели локтевой, лучевой, большой и малой берцовых костей. Последним фрагментом конечности будет аутоплодий, из которого образуются хрящевые модели костей кисти, стопы и фаланг пальцев.
Каждый этап дифференцировки клеток в процессе формирования парных конечностей позвоночных сопровождается либо активацией, либо подавлением экспрессии определенных генов.
Основные направления клеточной дифференцировки в процессе становления суставов в онтогенезе
Все компоненты сустава имеют общим источником развития скелетогенную мезенхиму сложного генеза, которая дифференцируется в нескольких направлениях, формируя соединительную, хрящевую и костную ткани. Основная функциональная задача структур с опорной функцией в процессе развития - это приобретение прочности, упругости, способности к обратимой деформации для преодоления биомеханических нагрузок при компрессии, растяжении и фрикции. Эта задача осуществляется волокнами и основным веществом матрикса, продуцентами которых являются клетки упомянутых тканей.
Приобретение суставными структурами необходимых биомеханических свойств идет различными путями и сопряжено со специфическими процессами, такими как фибробласто- и фибрилогенез, хондрогенез, остеогенез, а также синовиогенез.
Начальным этапом всех перечисленных направлений дифференцировок служит формирование волокнистого коллагенового каркаса и основного вещества, представленного протеогликанами и гликопротеинами. Такой соединительнотканный скелет присущ низшим позвоночным, и его формирование - это облигатный этап в развитии скелета всех высших позвоночных и человека. Последующие дифференцировки имеют существенные отличия.
Так, в процессе фибробласто- и фибрилогенеза происходит преимущественное развитие прочных разнонаправленных, но всегда определенным образом ориентированных волокнистых конструкций, обеспечивающих этим структурам взаимное смещение, растяжение, а также упругость и обратимость возникающей деформации. Так построены капсулы, связки, СО суставов.
Специфика развития матрикса в процессе хондрогенеза заключается в его гипергидратации, которая возможна за счет полианионных свойств протеогликанов и заключается в способности как удерживать, так и отдавать (возвращать) интерстициальную воду. Такой принцип приобретения необходимых биомеханических свойств присущ хрящевым компонентам скелета высших позвоночных и человека, а также группе хрящеподобных, хондроидных тканей скелетов низших позвоночных и хрящей внескелетных образований у высших (например, хондроидная ткань сердца и др.). Одной из самых ранних закладок в эмбриональном развитии скелета и наиболее древней филогенетически - хорде (chorda dorsales) - присущи те же биомеханические характеристики.
Прочностные свойства формирующейся костной ткани в процессе остеогенеза обеспечиваются минерализацией органической основы матрикса с участием неорганических соединений, в первую очередь солей кальция и фосфора, в результате чего формируются твердые и прочные костные пластины, образующие трабекулы. Такой же способ приобретения способности противостоять биомеханическим нагрузкам присущ экзоскелету древних позвоночных.
Органоспецифичной дифференцировкой, характерной только для синовиальных суставов, является синовиогенез. Внутренний слой капсулы синовиального сустава - СО - характеризуется структурными и функциональными особенностями покровного слоя, который непосредственно контактирует с суставной полостью, поддерживает ее существование и принимает участие в образовании СЖ.
Ранние этапы формирования сегментов скелета
Прехондральная мезенхима
Как известно, формирование скелета у всех позвоночных начинается с хрящевых моделей будущих образований костных сегментов. В эмбриональном онтогенезе в скелетогенной мезенхиме содержатся группы клеток, которые располагаются рыхло, не образуют агрегатов, имеют овальное или округлое ядро и органеллы цитоплазмы, присущие всем активно пролиферирующим и секретирующим клеткам. Совокупность таких клеток принято называть прехондральной мезенхимой.
Подобные клетки рассматриваются как плюрипотентные с еще не определившимся направлением дифференцировки. Полагают, что среди них уже существуют субпопуляции клеток прехондральной мезенхимы скелетогенного зачатка, детерминированных для последующего развития или только в хрящ, или только в соединительную ткань. Коммитированность этих клеток морфологически еще не выражена, они находятся в состоянии скрытой, или протодифференциации, а следовательно, их потенции к дифференцировке пока еще не реализованы.
Хрящевая бластема
Клетки одной из субпопуляций прехондральной мезенхимы обнаруживают тенденцию к конденсации: образуются плотные агрегаты клеток. Вслед за этим основное вещество, окружающее агрегаты клеток, приобретает тинкториальные свойства, присущие хрящу. Процессы хондрогенной дифференцировки, предшествующие ее морфологическому выражению, хорошо изучены на молекулярном и надмолекулярном уровнях.
Известно, что процесс конденсации клеток в бластеме является критической стадией в развитии скелета. Мутантные гены, определяющие возникновение пороков развития суставов, проявляют свое действие именно на стадии конденсации клеток.
Затем агрегированные хондрогенные клетки в мезенхимном зачатке конечности становятся морфологически отличимыми от клеток, развивающихся в направлении фибробластогенеза. Такие клетки принято называть хондробластами.
Процессы дифференцировки хрящевой бластемы, предшествующие кавитации
Интерстициальный рост хрящевой бластемы. Дальнейший рост и дифференцировка хрящевых закладок связаны с пролиферацией клеток бластемы и активной секрецией ими компонентов матрикса хряща. Структурные изменения в хондробластах связаны с характером экспрессии генов, программирующих синтез коллагенов: клетки полностью переключаются с синтеза коллагена I типа на синтез коллагена II типа. Почти одновременно в этих клетках начинается синтез стержневого белка агрекана. Это было показано на материале ранних стадий развития куриных эмбрионов с применением иммуногистохимических методов, когда одновременно использовались сыворотки к коллагенам I и II типа и к агрекану.
Формирование перихондра (надхрящницы). Перихондр формируется клетками окружающей хрящевую бластему околохрящевой перихондральной мезенхимы.
Наружный слой перихондра формируется клетками фибробластического типа, продуцирующими соединительнотканный матрикс, и внутренний, прилежащий к хрящу слой, клетки которого на всех последующих этапах развития сохраняют способность к хондрогенной дифференцировке. Именно за счет этих клеток осуществляется аппозиционный рост хряща, который заключается в нарастании новых масс ткани по периферии.
Фрагментация бластемы, формирование интерзоны. Длительное время конкретные механизмы фрагментации бластемы оставались неясными. В литературе обсуждались разные возможные механизмы этого процесса. Согласно существующим представлениям, фрагментация происходит благодаря внедрению в определенные участки хрящевой бластемы клеток перихондральной мезенхимы, которые коммитированны в направлении фибробластогенеза. Участки мезенхимы между хрящевыми фрагментами получили название «интерзон».
Рост и дифференцировка хрящевых моделей сочленяющихся костей
Формирование и готовность к функции основных компонентов внутренней среды сустава значительно опережают по времени завершение формирования костных сегментов сочленения. Будущие костные сегменты представлены провизорными гиалиновыми хрящами, осуществляющими рост хрящевой модели в длину и ширину.
Интерстициальный и аппозиционный рост провизорных хрящей
Рост хрящевых моделей костей, а именно увеличение их массы и объема осуществляются вследствие таких процессов, как увеличение числа и размеров хрящевых клеток и накопления продуцируемого клетками матрикса. В совокупности эти процессы обеспечивают интерстициалъный (внутритканевой) рост хряща, происходящий без нарушения его внутренней структуры. Способность к интерстициальному росту - уникальная особенность хрящевой ткани.
Вместе с тем хрящу, как и кости, свойствен и другой способ роста - наращивание массы ткани за счет продукции хондрогенных клеток перихондра (надхрящницы) на поверхности хряща. Такой способ получил название аппозиционного роста.
Рост провизорных хрящевых моделей длинных трубчатых костей конечностей осуществляется сочетанным проявлением упомянутых способов. Оно неодинаково у позвоночных различных таксономических групп и у человека в разные периоды формирования конечностей. Рост в длину всегда имеет в своей основе интерстициальный механизм. Кинетика этого процесса изучена методами с использованием радиоактивной метки на зародышах млекопитающих и на куриных эмбрионах. Интенсивность роста весьма значительна. Так, у крысы проксимальный конец большеберцовой кости увеличивается в длину на 130 мкм в день; у человека та же кость удлиняется на 30 мкм в день.
Рост в ширину осуществляется как интерстициально, так и аппозиционно. Аппозиционный механизм включается на более поздних стадиях развития. Клетки внутреннего слоя надхрящницы, сохраняющие хондрогенные потенции, мигрируют к периферическим участкам метаэпифизарной пластинки и дают начало новым клеткам, которые располагаются продольно, образуя добавочные колонки. Тем самым в формирующейся кости увеличивается ее поперечный размер. Подобным способом увеличивается и поперечный размер эпифиза. Скорость роста хрящевой модели в ширину (или поперечного роста) определяется как V5-V10 от скорости роста в длину.
Рост и дифференцировка хрящевых эпифизов
Для изучения закономерностей развития суставов наибольший интерес представляют процессы формирования эпифизарных концов хрящевых моделей. В росте хрящевых эпифизов принято различать три стадии:
- предкавитационную (раннюю);
- предоссификационную;
- постоссификационную.
Предкавитационная (ранняя) стадия уже описана выше. Она включает процессы формирования хрящевой бластемы, ее фрагментацию, образование перихондра и интерзоны между соседними хрящевыми фрагментами. В интерзоне происходит процесс кавитации. На этой стадии в клетках хрящевой модели эпифиза выявляются митозы и происходит накопление массы матрикса и увеличение объема клеток. Это дает основание говорить об интерстициалъном росте хряща. Однако, по признанию многих исследователей, ведущим процессом на этой стадии является аппозиционный рост на поверхности хрящевых фрагментов.
В начале следующей второй - предоссификационной стадии, которая охватывает период после начала кавитации до начала образования очагов оссификации в эпифизе, формируется суставной гиалиновый хрящ, который покрывает поверхности будущих костей. У суставного хряща перихондр отсутствует.
В глубоких слоях хрящевого эпифиза наблюдаются митозы, а в периферийных участках хрящевой модели идет аппозиционный рост за счет клеток перихондра. Перихондр формируется на ранних стадиях образования хрящевых моделей, состоит из двух слоев: 1) наружного волокнистого, богатого сосудами; 2) внутреннего, обращенного к хрящу, - хондрогенного. Хондрогенные клетки являлись источником аппозиционного роста самого хряща, однако на последующих этапах генетическая программа этих клеток реализуется в направлении дифференцировки остеогенных клеток - остеобластов, секреторная деятельность которых приводит к образованию матриксных структур кости. В дальнейшем остеогенные клетки перихондра, теперь уже периоста (надкостницы), проникают вглубь в центр эпифиза, где формируется новый очаг оссификации.
Эпифизарный очаг оссификации формируется за счет процессов, аналогичных таковым в диафизе хрящевой модели, т.е. процессов энхондральной оссификации. В центре эпифиза зрелые хондроциты гипертрофируются, матрикс хряща кальцифицируется, клетки гибнут, и образуется полость. В гибнущий хрящ со стороны перихондра внедряются сосуды, сопровождающие их перициты и остеогенные клетки - остеобласты, которые продуцируют вещества матриксных структур. Так формируется молодая ретикулофиброзная кость - первичный очаг оссификации эпифиза.
Третья постоссификационная стадия развития хрящевых эпифизов - период от формирования эпифизарного центра оссификации до полной замены хряща костью - характеризуется замедлением интенсивности продольного и поперечного роста хряща. Вместе с тем интерстициальный рост продолжается, и осуществляется аппозиция со стороны перихондра. Очаг оссификации распространяется от центра эпифиза к периферии. Участок кости, контактирующий с хрящевой пластинкой, покрывающей суставную поверхность, именуется субхондральной костью.
После того как эпифиз сочленяющейся кости достигнет своего дефинитивного размера, пролиферация в хряще и замещение его костью прекращаются, но в дальнейшей постнатальной жизни в норме толщина суставного хряща (иная в различных суставах и в разных участках суставной поверхности) остается постоянной.
Гистогенетические процессы при формировании эпифизов
Наблюдения за развитием хрящевой бластемы в условиях in vitro (в культуре ткани и при культивировании на хорионаллан-тоисе куриного эмбриона) показали, что наряду с цитогенетическими дифференцировками, описанными выше, бластема обладает и морфогенетическими потенциями, т.е. обеспечивает условия, при которых конкретный фрагмент в процессе роста приобретает ту форму, свойственную ему при развитии in vivo.
На ранней стадии формирования хрящевых эпифизов, как это было показано на примере куриных эмбрионов (стадии 22-24), даже удаление части бластемы компенсируется за счет пролифера-тивных возможностей ее клеток. В итоге формируется нормальный скелет данного участка конечности.
Гистогенетические процессы при формировании хрящевых моделей костей сочетаются с органогенетическими, ибо показано, что именно на хрящевой стадии закладки кости приобретают характерные анатомические особенности. В основе изменения формы хрящевых закладок лежит ряд факторов. Богатый водой матрикс хрящевой ткани обладает вязкоупругими свойствами и способностью изменять свой объем под механическим воздействием других структурных элементов.
Перихондр способен сдерживать увеличение объема хряща, что наиболее выражено в диафизарных частях хрящевой закладки и в меньшей степени - в эпифизах, где давление со стороны растущего хряща превалирует. Коррекция формирования растущих эпифизов связана с их контактами с эпифизами смежных сегментов хрящевых моделей. Определенное значение имеют генетически детерминированные различия в сроках дифференцировки клеток в разных участках хрящевой модели, в частности так называемых пролиферативных центрах.
Остеогенетические процессы в развитии сочленяющихся костей
Оссификация хрящей эпифизов начинается с формирования центрального ядра за счет гипертрофии и гибели хондроцитов, отложения кальциевых солей в матриксе и проникновения в участки деградированного хряща сосудов и остеогенных клеток со стороны перихондра.
Описание процессов окостенения хрящевых моделей сочленяющихся костей, по-видимому, целесообразно предварить четкой договоренностью о термине и об соотношениях определяемых им процессов. Образование минерального компонента костной ткани является результатом процесса минерализации. Понятие минерализации не следует смешивать с понятиями кальцификации и оссификации. Кальцификация - более общее понятие, обозначающее отложение разнообразных кальциевых солей обычно в различных тканях, иногда носящее патологический характер. Процесс кальцификации широко представлен у представителей обоих ветвей родословного древа животного мира. Ведущий компонент в скелете вторичноротых (в ряду позвоночных) - фосфат кальция, у первичноротых - карбонат кальция.
Минерализация - частный случай кальцификации - отложение в органическом матриксе костной ткани кальций-фосфорных солей в кристаллической форме (аналогичный процесс происходит и в ткани зубов). Что же касается оссификации, то это понятие объединяет весь процесс формирования костной ткани путем дифференциации предшествующей ей мезенхимы или замещения хрящевой ткани; этот специфический процесс включает в себя минерализацию в качестве заключительного этапа.
Минерализация рассматривается как двухэтапный процесс. Первый этап - образование и накопление достаточной концентрации кальций-фосфорных соединений, которые протекают в так называемых матричных пузырьках. Матричные пузырьки - это покрытые мембраной экстрацеллюлярные частицы диаметром от 30 до 200 нм, избирательно обнаруживаемые в очагах начинающейся кальцификации. Пузырьки развиваются путем образования выпячиваний в специализированных, обращенных в сторону кальцифицируемого матрикса участках цитоплазматической мембраны, ответственных за кальцификацию клеток. Сформировавшиеся выпячивания отделяются от клеток и превращаются в пузырьки. Имеются данные о существовании связей между этими пузырьками и коллагеновыми волокнами.
В Накоплении кальций-фосфорных соединений в пузырьках участвуют содержащиеся в них ферменты, обеспечивающие накопление ионов фосфата, фосфатазы. В накоплении ионов кальция участвуют также связывающие кальций молекулы, такие как аннексии V (анкорин СII) и аннексии - А1 (липокортин I), члены семейства аннексинов и белков, образующие кальциевые каналы в клеточных мембранах; белки кальбиндин и калъпактин и соединение липидной природы фосфатидил серина.
При достижении в жидкости внутри матричных пузырьков необходимой концентрации гидроксиапатита, образующегося в результате химической реакции между ионами кальция и ионами ортофосфата, начинается собственно кристаллизация, происходящая вблизи внутренней поверхности их мембраны. Кристаллизация не является химической реакцией, а представляет собой фазовую трансформацию, аналогичную трансформации воды в лед. Начало кристаллизации именуется нуклеацией. Она заключается в образовании мельчайших кристаллических ядер будущих кристаллов и происходит в соприкосновении с органическими макромолекулами, т.е. является гетерогенной.
Второй этап минерализации костной ткани начинается после того, как первичные кристаллы гидроксиапатита (ядра) освободятся из матричных пузырьков и соприкоснутся с внеклеточной (интерстициальной) жидкостью. В этой жидкости концентрация Са 2+ и РО 3- 4 достаточна для того, чтобы обеспечить дальнейший рост кристаллов, протекающий по аппозиционному типу, путем последовательного присоединения молекул из тканевой жидкости. Рост первичных кристаллов происходит в основном в пробелах между концами макромолекул коллагена I типа внутри коллагеновых фибрилл. По мере роста кристаллы захватывают все свободные промежутки между соседними фибриллами в коллагеновых волокнах, и в процесс вовлекаются неколлагеновые белки. Время, необходимое для этого, невелико - оно измеряется несколькими часами от момента начала кристаллизации.
Процессы оссификации
При описании сложных процессов замены хряща костью в хрящевых моделях целесообразно подчеркнуть сочетанность процессов в гибнущем и кальцифицирующемся хряще и продуктивных процессов, осуществляемых внедрившимися в хрящ остеобластами. В связи с этим необходимо остановиться на ряде следующих закономерностей. К факторам, непосредственно вызывающим кальцификацию и гибель хряща в очаге окостенения, относят:
- способность гипертрофированных хондроцитов секретировать щелочную фосфатазу, присутствие которой обеспечивает изменение соотношения ортофосфатов и пирофосфатов, необходимых для отложения гидроксиапатита;
- активное выделение хондроцитами матриксных пузырьков;
- врастание капилляров при распаде кальцифицированного матрикса.
Время и последовательность выявления этих процессов определяются генетической программой. Внедряющиеся кровеносные капилляры способствуют усилению оксигенизации процессов метаболизма. Последнее необходимо для существования кости. Тот же фактор является определяющим для изменения характера дифференцировки во внутреннем слое перихондра, где плюрипотентные клетки, являвшиеся источником хондрогенных клеток, теперь уже становятся остеогенными, т.е. строящими костное вещество.
После того как эпифиз сочленяющейся кости достигнет своего дефинитивного размера, пролиферация в хряще и замещение его костью прекращаются. В дальнейшей постнатальной жизни толщина суставного хряща в норме остается постоянной, однако в различных суставах и разных участках суставной поверхности она различна.
Оссификация и кровоснабжение эпифизов развивающихся костей
Оссификация. Та часть хрящевого эпифиза, которая входит в сочленение, содержит следующие компоненты: эпифизарный хрящ, хондроциты которого находятся на различных стадиях жизненного цикла; кальцифицирующееся основное вещество хряща; костный компонент, представленный очагом (центром) оссификации данного эпифиза; узкий участок эпифизарного гиалинового хряща, открытого в суставную полость. Границей между собственно эпифизом и диафизарной частью кости является эпифизарная (или метаэпифизарная) пластинка хрящевой ткани, за счет которой осуществляется рост всей кости в длину, завершающийся лишь в постнатальном онтогенезе".
Процессы кальцификации матрикса и гибели хондроцитов в хряще эпифизов распространяются во все стороны от очага оссификации. Молодая кость - продукт секреции веществ остеобластами - располагается на остатках хряща. В условиях усиливающихся биомеханических нагрузок расположение трабекул в формирующейся кости приобретает упорядоченность в соответствии с направлением сил, действующих при движениях. Образуется центральная губчатая часть эпифиза. В сердцевине каждой костной трабекулы сохраняется участок кальцифицированного хряща. В ячеях, образованных трабекулами губчатого вещества, формируется миелоидная ткань.
Периферия молодой эпифизарной кости приобретает свойства компактной кости с формированием в ней первичных гаверсовых систем. Этот участок эпифизарной кости непосредственно подлежит суставному хрящу и называется субхондральной костью.
Кровоснабжение эпифизов развивающейся кости. Усиленная потребность в кровоснабжении формирующейся кости в костных компонентах сустава удовлетворяется поступлением в хрящевую модель кровеносных сосудов из ряда источников. Один из них, включающийся на самых ранних стадиях, - это сосуды периостальной почки, прорывающиеся внутрь хряща от перихондра (периоста). Эти сосуды получили название питающих артерий и вен.
Другой источник - сосуды метафизарной части формирующейся кости. На более поздних стадиях развития сосуды из периферической части эпифизарной пластинки проникают к перихондру (теперь периосту), а затем вместе с сосудами периоста - в эпифиз, где анастомозируют с питающими сосудами. Хрящ, сохраняющийся длительное время в эпифизарной пластинке, может получать питание от сосудов кости как с диафизарной, так и с эпифизарной ее стороны.
Общая характеристика гистогенетических процессов становления сустава
Для характеристики процессов органогенеза суставов во второй половине антенатального периода развития наибольший интерес имеют данные о составе СЖ у плодов. Из коленных и других крупных суставов плодов человека 4,5-7 месяцев внутриутробной жизни с помощью меланжера для крови или методом получения реплик с поверхности суставного хряща получали несколько капель синовии. В них определялось значительное количество клеток (до 100).
Выяснилось, что в синовии плодов содержатся клетки формирующейся СО и клетки крови. В составе первой преобладающей группы отчетливо идентифицируются молодые кроющие клетки - синовиобласты и фибробласты. Последние, как известно, не свойственны синовии взрослых людей и животных. В жидкой части синовии обнаруживается присутствие гиалуронана.
Становление суставов в онтогенезе представляет собой скоординированную систему последовательно протекающих биохимических взаимодействий, обеспечивающую изменения архитектоники и обмена в определенных отделах хрящевого скелета. Последние создают условия для приложения биомеханических факторов.
По выражению В.К. Hall, при артрогенезе возникает «генетически заданный и прогнозируемый временной каскад», подверженный постоянному воздействию многих регуляторных механизмов.
Возможное на основе существующих данных сопоставление морфологии развивающихся структур сустава с «каскадом морфогенетических процессов» и их биохимической характеристикой представлено в следующей сводной таблице.
| Морфологически выявляемые структуры и процессы | Морфогенетические закономерности | Основные синтезы |
|---|---|---|
| 1 | 2 | 3 |
| I. Скелетогенная мезенхима - совокупность свободно расположенных, морфологически не различающихся пролиферирующих клеток | I. Плюрипотентные пролиферирующие клетки, еще не коммитированные в отношении развития в одном определенном направлении, подвергающиеся клеточной дифференцировки и индукционному воздействию производных других эмбриональных закладок | I. Продукция и секреция коллагена I типа, фибронектина и гиалуронана |
| II. Перихондральная мезенхима - конденсация клеток вследствие их миграции; образование агрегатов взаимодействующих клеток | II. Выявление популяции клеток прехондробластов, коммитированных в хондрогеном направлении клеточной дифференцировки. Факторы детерминации: как и реализация генетической программы; индукционное воздействие со стороны производных других эмбриональных закладок | II. Те же синтезы |
| III. Хрящевая бластема. Интерстициальный рост бластем как результат образования и накопления вещества экстрацеллюлярного матрикса; приобретение клетками фенотипических черт хондробласта; и начало формирования перихондра; обособление хрящевой бластемы от клеток парахондральной мезенхимы | III. Хондрогенная дифференцировка: приобретение клетками сферической формы, удлинение времени жизненного цикла (до периода интерфазы), межклеточные взаимодействия в цитоплазме. Активизация процесса дифференцировки органоидов | III. Начало синтеза и секреции хрящевыми клетками коллагена II типа и сульфатированных гликозаминогликанов, угнетение синтезов гиалуронана и фибронектина |
| IV. Фрагментация бластемы. Обособление в бластеме хрящевых сегментов; образование между сегментами: вначале однослойной, а затем трехслойной интерзоны; приобретение клетками бластемы фенотипических черт хондроцита | IV. В хрящевых клетках - активизация развития цитоплазматических структур, ответственных за синтез и секрецию веществ, формирующих экстрацеллюлярный метрике. Накопление волокон и основного вещества матрикса, приводящее к рассредоточению клеток. Выявление вязкоупругих свойств матрикса | IV. Синтез и секреция хрящевыми клетками коллагена II типа и сульфатированных гликозаминогликанов |
| V. Кавитация - образование суставной полости. Образование в интерзоне ряда щелевидных пространств и их слияние в одну щелевидную полость, формирование по периферии интерзоны популяции клеток - будущей синовиальной оболочки | V. Способы (механизмы) кавитации - это совокупность процессов, таких, как: генетически запрограммированная гибель части хондроцитов в хрящевой ткани интерзоны; дальнейшее накопление матрикса, способствующая рассредоточению клеток; продукция фибробластическими клетками интерзоны гиалуронана. | V. В хондроцитах - синтез коллагена II типа и сульфатированных гликозаминогликанов. В синовиобластах и клетках парахондральной мезенхимы - синтез коллагена I типа и гиалуронана |
| VI. Хрящевые модели сочленяющихся фрагментов скелета, их интерстициальный и оппозиционный рост. Формирование эпифизов. Перемещение суставных концов сочленяющихся фрагментов скелета по отношению друг к другу - в начале в связи со спонтанной, а затем инициированной нервной импульсацией,обеспечивающей сокращения мускулатуры | VI. Продолжение пролиферативных и продуктивных процессов в хондроцитах: увеличение числа и размеров клеток, накопление веществ матрикса. Дифференцировка клеток и матрикса синовиальной оболочки | VI. Те же синтезы в клетках ткани хряща и формирующейся синовиальной оболочки |
| VII. Периостальная манжетка в диафизах хрящевых моделей. Формирование эпифизов, органогенетические процессы. | VII. Дифференцировка клеток перихондра: изменение фенотипа его клеток, формирование остеобластов | VII. Те же синтезы в клетках ткани хрящевой модели. Синтез коллагена I типа и гиалуронана в клетках формирующейся СО |
| VIII. Кальцификация и гибель клеток в диафизах хрящевых моделей костей. Проникновение в хрящ из периоста остеобластов и сосудов - начальные процессы парахондральной оссификации | VIII. Отложение на коллагеновых фибриллах матрикса хряща гидроксиапатита, рост первичных ядер отложений кальция. Остеогенетическая деятельность остеобластов, в хряще и продукция ими костного вещества | VIII. В хондроцитах продолжение синтеза коллагена II типа и сульфатированных гликозаминогликанов; в хондроцитах энергетический обмен по типу гликолиза. Отложение гидроксиапатита. В остеобластах - синтез коллагена I и III типов и гиа-луронана. Энергетический обмен окислительного типа с постепенным его усилением по мере оксигенизации формирующейся костной ткани |
| IX. Формирование кости в диафизах (энхондральная оссификация); оссификация эпифизов. Формирование в поверхностной зоне в эпифизах пластинчатой кости - субхондральной костной пластинки. Образование в ячеях трабекул губчатой кости миелоидной ткани костного мозга | IX. Кальцификация и гибель хрящевой ткани. Продукция и секреция остеобластами костного вещества, упорядочение расположения костных пластинок в связи с формированием капиллярного звена васкуляризации. Дифференцировка части клеток в клетки миелоидной ткани костного мозга. Дифференцировка структур СО. Появление в суставной полости первичной синовиальной жидкости | IX. Сохранение свойственного хрящу синтезов коллагена II типа и сульфатированных гликозаминогликанов, а также энергетического обмена по типу - гликолиза. Усиление энергетического обмена окислительного типа. Появление клеток миелоидной ткани, проникающих в кость вместе с сосудами |
Сустав — это подвижное сочленение двух или более костей скелета.
Суставы объединяют кости скелета в единое целое. Двигаться человеку помогают более 180 различных суставов. Вместе с костями и связками их относят к пассивной части двигательного аппарата. Суставы можно сравнить с шарнирами, в задачу которых входит обеспечение плавного скольжения костей относительно друг друга. При их отсутствии кости будутпросто тереться друг о друга, постепенно разрушаясь, что является очень болезненным и опасным процессом. В организме человека суставы играют тройную роль: они содействуют сохранению положения тела, участвуют в перемещении частей тела относительно друг друга и являются органами локомоции (передвижения) тела в пространстве.
Основными элементами, которые имеются во всех так называемых истинных суставах, являются:
- суставные поверхности (концы) соединяющихся костей;
- суставная капсула;
- суставная полость.
Полость сустава заполняет синовиальная жидкость, которая является своеобразной смазкой и способствует свободному движению суставных концов.
По числу суставных поверхностей различают:
- простой сустав, имеющий только 2 суставные поверхности, например межфаланговые суставы;
- сложный сустав, имеющий более двух сочленяющихся поверхностей, например локтевой сустав. Сложный сустав состоит из нескольких простых сочленений, в которых движения могут совершаться отдельно;
- комплексный сустав, содержащий внутрисуставной хрящ, который разделяет сустав на 2 камеры (двухкамерный сустав).
Классификацию суставов проводят по следующим принципам:
- по числу суставных поверхностей;
- по форме суставных поверхностей;
- по функции.
Суставная поверхность кости образована гиалиновым (реже волокнистым) суставным хрящом. Суставные хрящи представляют собой ткань, наполненную жидкостью. Поверхность хряща ровная, крепкая и эластичная, способна хорошо впитывать и выделять жидкость. Толщина суставного хряща в среднем составляет 0,2-0,5 миллиметра.
Суставная капсула образована соединительной тканью. Она окружает сочленяющиеся концы костей и на суставных поверхностях переходит в надкостницу. Капсула имеет толстую наружную волокнистую фибринозную мембрану и внутреннюю тонкую синовиальную мембрану, которая выделяет в полость сустава синовиальную жидкость. Связки и сухожилия мышц укрепляют капсулу и способствуют движению сустава по определенным направлениям.
К вспомогательным образованиям сустава относят внутрисуставные хрящи, диски, мениски, губы и внутрикапсульные связки. Кровоснабжение сустава осуществляется из широко анастомозирующей (разветвленной) суставной артериальной сети, образованной 3-8 артериями. Иннервация (снабжение нервами) сустава осуществляется нервной сетью, образованной симпатическими и спинномозговыми нервами. Все суставные элементы, кроме гиалинового хряща, имеют иннервацию. В них обнаруживаются значительные количества нервных окончаний, осуществляющих болевое восприятие, вследствие этого они могут стать источником боли.
Суставы обычно делят на 3 группы:
- синартрозы - неподвижные (фиксированные);
- амфиартрозы (полусуставы) - частично подвижные;
- диартрозы (истинные суставы) - подвижные. Большинство суставов относится к подвижным сочленениям.
По данным Всемирной организации здравоохранения от болей в суставах страдает каждый 7-й житель планеты. В возрасте от 40 до 70 лет заболевания суставов наблюдаются у 50% людей и у 90% людей старше 70 лет.
Синовиальный сустав - это сустав, в котором окончания костей сходятся в суставной сумке. К таковым относятся большинство суставов человека, в том числе несущие - коленный и тазобедренный суставы.
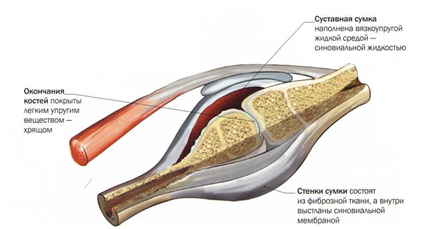
Суставы разделяют на простые и сложные. В образовании простых участвуют 2 кости, сложных - более 2 костей. Если в движении участвуют несколько самостоятельных суставов, как у нижней челюсти при жевании, такие суставы называются комбинированными. Комбинированный сустав представляет собой сочетание нескольких изолированных друг от друга суставов, расположенных отдельно, но функционирующих вместе. Таковы, например, оба височно-нижнечелюстных сустава, проксимальный и дистальный лучелоктевые суставы и другие.
По форме суставные поверхности напоминают отрезки поверхностей геометрических тел: цилиндра, эллипса, шара. В зависимости от этого различают цилиндрический, эллипсовидный и шаровидный суставы.
Форма суставных поверхностей определяет объем и направление движений вокруг 3 осей: сагиттальной (проходит по направлению спереди назад), фронтальной (проходит параллельно плоскости опоры) и вертикальной (перпендикулярна к плоскости опоры).
Круговое движение - это последовательное движение вокруг всех осей. При этом один конец кости описывает круг, а вся кость - фигуру конуса. Возможны и скользящие движения суставных поверхностей, а также удаление их друг от друга, как это, например, наблюдается при растягивании пальцев. Функция сустава определяется количеством осей, вокруг которых совершаются движения.
Различают следующие основные виды движений в суставах:
- движение вокруг фронтальной оси - сгибание и разгибание;
- движения вокруг сагиттальной оси - приведение и отведение движения вокруг вертикальной оси, то есть вращение: кнутри (пронация) и кнаружи (супинация).
Кисть человека содержит: 27 костей, 29 суставов, 123 связки, 48 нервов и 30 названных артерий. В течение жизни мы совершаем движения пальцами миллионы раз. Движение кисти и пальцев обеспечивают 34 мышцы, только при движении большого пальца задействуются 9 разных мышц.
Он самый подвижный у человека и образован головкой плечевой кости и суставной впадиной лопатки.
Суставная поверхность лопатки окружена кольцом фиброзного хряща - так называемой суставной губой. Через полость сустава проходит сухожилие длинной головки двуглавой мышцы плеча. Плечевой сустав укрепляет мощная клювовидноплечевая связка и окружающие мышцы - дельтовидная, подлопаточная, над- и подостные, большая и малая круглые. В движениях плеча принимают участие также большая грудная и широчайшая мышцы спины.
Синовиальная оболочка тонкой суставной капсулы образует 2 внесуставных заворота - сухожилия двуглавой мышцы плеча и подлопаточной мышцы. В кровоснабжении этого сустава принимают участие передняя и задняя артерии, огибающие плечевую кость, и грудоакромиальная артерия, венозный отток осуществляется в подмышечную вену. Отток лимфы происходит в лимфатические узлы подмышечной области. Плечевой сустав иннервируется ветвями подмышечного нерва.

- плечевая кость;
- лопатка;
- ключица;
- суставная капсула;
- складки суставной капсулы;
- акромио-ключичный сустав.
В плечевом суставе возможны движения вокруг 3 осей. Сгибание ограничивается акромиальным и клювовидным отростками лопатки, а также клювовидно-плечевой связкой, разгибание-акромионом, клювовидно-плечевой связкой и капсулой сустава. Отведение в суставе возможно до 90°, а с участием пояса верхних конечностей (при включении грудино-ключичного сустава) - до 180°. Прекращается отведение в момент упора большого бугра плечевой кости в клювовидно-акромиальную связку. Шаровидная форма суставной поверхности позволяет человеку поднимать руку, отводить ее назад, вращать плечо вместе с предплечьем, кистью внутрь и наружу. Такое разнообразие движений руки стало решающим шагом в процессе эволюции человека. Плечевой пояс и плечевой сустав в большинстве случаев функционируют как единое функциональное образование.
Он самый мощный и сильно нагружаемый сустав в организме человека и образован вертлужной впадиной тазовой кости и головкой бедренной кости. Тазобедренный сустав укреплен внутрисуставной связкой головки бедренной кисти, а также поперечной связкой вертлужной впадины, охватывающей шейку бедренной кости. Снаружи в капсулу вплетаются мощная подвздошно-бедренная, лобково-бедренная и седалищно-бедренная связки.

Кровоснабжение этого сустава осуществляется через артерии, огибающие бедренную кость, ветвями запирательной и (непостоянно) ветвями верхней прободающей, ягодичных и внутренней половой артерий. Отток крови происходит по венам, окружающим бедренную кость, в бедренную вену и через запирательные вены в подвздошную вену. Лимфоотток осуществляется в лимфатические узлы, расположенные вокруг наружных и внутренних подвздошных сосудов. Тазобедренный сустав иннервируется бедренным, запирательным, седалищным, верхним и нижним ягодичными и половыми нервами.
Тазобедренный сустав - разновидность шаровидного сустава. В нем возможны движения вокруг фронтальной оси (сгибание и разгибание), вокруг сагиттальной оси (отведение и приведение) и вокруг вертикальной оси (наружная и внутренняя ротация).
Данный сустав испытывает большую нагрузку, поэтому неудивительно, что поражения его занимают первое место в общей патологии суставного аппарата.
Один из крупных и сложно устроенных суставов человека. Его образуют 3 кости: бедренная, большеберцовая и малоберцовая. Стабильность коленному суставу обеспечивают внутри- и внесуставные связки. Внесуставными связками сустава являются малоберцовая и большеберцовая коллатеральные связки, косая и дугообразная подколенные связки, связка надколенника, медиальная и латеральная поддерживающие связки надколенника. К внутрисуставным связкам относятся передняя и задняя крестообразные связки.
Сустав имеет много вспомогательных элементов, таких как мениски, внутрисуставные связки, синовиальные складки, синовиальные сумки. В каждом коленном суставе имеются по 2 мениска - наружный и внутренний. Мениски имеют вид полулуний и выполняют амортизационную роль. К вспомогательным элементам этого сустава относятся синовиальные складки, которые образуются синовиальной мембраной капсулы. Коленный сустав также имеет несколько синовиальных сумок, часть из которых сообщается с полостью сустава.
Каждому приходилось восхищаться выступлениями спортивных гимнасток и артистов цирка. О людях, способных залезать в небольшие ящики и неестественно выгибаться, говорят, что у них гуттаперчевые суставы. Разумеется, это не так. Авторы «Оксфордского справочника органов тела» уверяют читателей, что «у таких людей суставы феноменально гибки» - в медицине это называется синдромом гипермобильности суставов.

- бедренная кость
- большеберцовая кость
- синовиальная жидкость
- внутренний и наружный мениски
- медиальная связка
- латеральная связка
- крестообразная связка
- надколенник
По форме сустав является мыщелковым суставом. В нем возможны движения вокруг 2 осей: фронтальной и вертикальной (при согнутом положении в суставе). Вокруг фронтальной оси происходит сгибание и разгибание, вокруг вертикальной оси - вращение.
Коленный сустав очень важен для передвижения человека. При каждом шаге за счет сгибания он дает возможность ноге шагнуть вперед без удара о землю. Иначе нога выносилась бы вперед за счет поднятия бедра.


